Содержание
Особенности вируса. Вирус кори относится к роду Morbillivirus, который включает наиболее агрессивные парамиксовирусы. Кроме вируса кори он объединяет возбудителей чумы собак (поражает широкий круг плотоядных), крупного рогатого скота, овец и коз (опасен также для свиней), морских млекопитающих (китов, дельфинов, тюленей), смертельной пневмонии лошадей (известны случаи фатального заражения людей). Первые два связаны с вирусом кори антигенным родством и не дают перекрестных реакций с другими парамиксовирусами. Они вызывают сходные заболевания, нередко с фатальным исходом. Достаточно вспомнить о собачьей чуме (англ. canine distemper), особенно опасной для щенков; она протекает с тяжелым диарейным синдромом, поражением центральной нервной системы и легких, убивая или оставляя тяжелые последствия.
Вирус кори не только крайне контагиозен, но и высоковирулентен: в отличие от других парамиксовирусов бессимптомного заражения практически не бывает. При внесении вируса в популяцию, лишенную эндемичной инфекции (она поддерживает коллективный иммунитет), эпидемия неизбежна, причем болеет почти 100% населения. Последствия подобных вспышек (они известны из истории освоения европейцами новых земель, прежде всего Америки) бывали катастрофичны. Яркий пример — вспышка кори, случившаяся в 1846 г. среди жителей Фарерских Островов после общения с иммигрантами из Европы. В течение 6 нед переболело около 4000 человек. Исключение составили всего 5 человек старше 60 лет, которые много лет назад пережили последнюю коревую эпидемию и благодаря этому приобрели иммунитет.
Вирус кори похож на другие парамиксовирусы. Тем не менее есть различия, которые, возможно, имеют отношение к патогенезу инфекции:
- суперкапсидные шипы содержат только гемагглютинин, но не обладают нейраминидазной активностью (это же характерно для респираторно-синцитиального вируса);
- спектр гемагглютинационной активности ограничен эритроцитами обезьян; вирус кори не агглютинирует эритроциты человека, кур, морских свинок, мышей и пр., которые агглютинируются другими парамиксовирусами;
- связывание вириона не зависит от сиаловых кислот, но основано на взаимодействии с CD46 и, возможно, с другими мембранными структурами;
- цитопатический эффект сопровождается формированием не только цитоплазматических, но и внутриядерных включений (хотя репликация, как и у других парамиксовирусов, происходит в цитоплазме);
- симпластообразующая активность проявляется не только in vitro, но и в тканях инфицированного организма;
- в перевиваемых клеточных культурах кроме острой может быть смоделирована и персистентная инфекция без продукции инфекционного вируса и цитопатогенного эффекта;
- кроме эпителиальных клеток вирус размножается в мононуклеарных фагоцитах, лимфоцитах и эндотелиальных клетках.
Вирус кори имеет единственный серотип, не претерпевший существенных изменений с момента открытия (первый штамм изолирован J.F. Enders и T.C. Peebles в 1954 г.). Эпитопные различия, регистрируемые между штаммами при помощи моноклональных антител, не влияют на антигенную монолитность вируса в реакциях протективного иммунитета. Единственным природным резервуаром вируса является человек, хотя искусственно могут быть инфицированы обезьяны, собаки, мыши и другие животные. Полноценную картину кори удается воспроизвести только на обезьянах.
Патогенез. Основной поражаемый контингент — дети 5—6 лет, но вакцинация изменила эпидемиологические параметры, в том числе возрастные приоритеты, сместив их на школьный возраст. Заражение обычно происходит воздушно-капельным путем. В течение нескольких дней вирус размножается в эпителиальных клетках дыхательного тракта, в макрофагах и дендритных клетках регионарных лимфатических узлов, откуда распространяется (первичная вирусемия) во все отделы лимфоидной и ретикуло-эндотелиальной ткани. Здесь продолжается его размножение, после чего следует вторая, более мощная волна вирусемии инфекции. Вирус переносится лейкоцитами, прежде всего моноцитами. Удивительно, но его не удается обнаружить в лимфоцитах. Возможно, инфицированные лимфоциты быстро удаляются из циркуляции, погибая в лимфоидной ткани, или просто не поддерживают репродукцию вируса.
Вирус проникает практически во все органы. Это объясняется заражением таких убиквитарных клеток, как макрофаги, эпителиоциты кожи и слизистых оболочек. Но центральным событием, безусловно, является инфекция эндотелиальных клеток мелких сосудов. Именно в зоне их поражения формируются воспалительные реакции, которые вносят решающий вклад в развитие болезни. Многоядерные гигантские клетки с внутриядерными включениями можно обнаружить в лимфоидной ткани (лимфатические узлы, селезенка, миндалины, аппендикс), эпидермисе, конъюнктиве, печени, почках, респираторном тракте, кишечнике. Инкубация составляет 9—14 дней, хотя иногда затягивается до трех недель. Продромальный период (его называют катаральным) совпадает с вторичной вирусемией и агрессивной диссеминацией вируса. Он продолжается 2—4 дня, напоминая ОРЗ (ринит, фарингит, кашель, конъюнктивит, умеренные признаки интоксикации). Уникальный признак — энантема («внутренняя сыпь») на слизистой оболочке щек. Это так называемые пятна Коплика—Филатова — очаги некроза с зоной воспалительной гиперемии диаметром 1—3 мм. Их появление позволяет поставить диагноз до «настоящей кори».
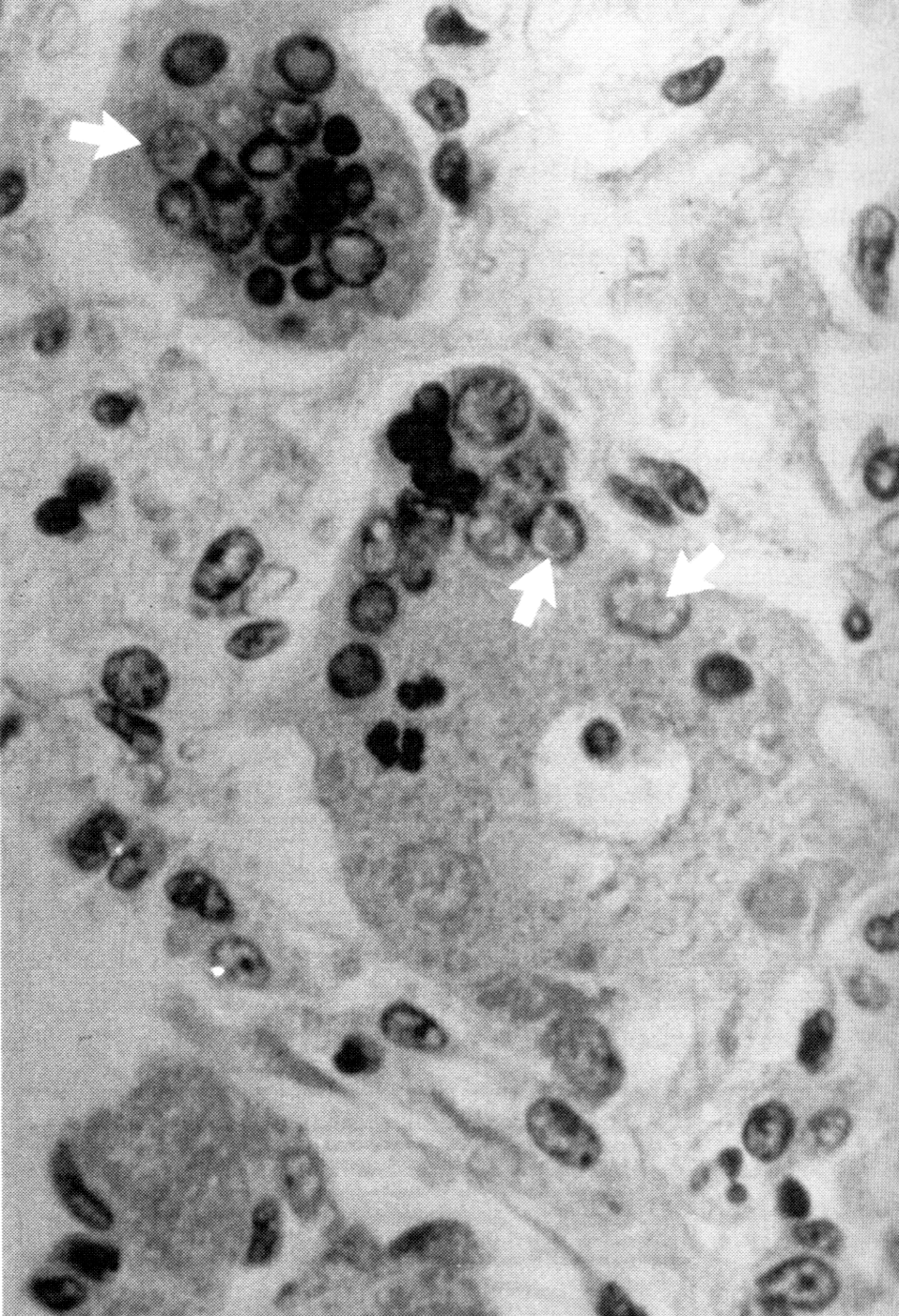
«Настоящую» корь (она следует за продромальным периодом) знаменуют папулезная, часто сливающаяся сыпь (экзантема) с характерной динамикой (начинаясь с шеи и лица, она распространяется на тело и конечности) и тяжелая интоксикация, прежде всего высокая температура. Экзантема представляет собой очаги воспаления эпителиальной ткани, которые формируются на основе иммунологически зависимого повреждения вирусинфицированного эндотелия мелких сосудов. У больных с дефектами клеточного иммунитета сыпь отсутствует, хотя другие проявления коревого синдрома протекают не менее тяжело и часто более опасны. На пике экзантемы состояние выглядит критическим, но через несколько дней обычно все заканчивается благополучно. Осложнения возникают редко. Чаще всего (5—15%) это вторичные бактериальные инфекции — средний отит, синуситы, бронхопневмония. К ним предрасполагает нарушение клиринговой функции респираторного тракта (повреждение мерцательного эпителия) и временное ослабление иммунитета, типичное для коревой инфекции. Следствием прямой вирусной агрессии может быть интерстициальная пневмония, для которой характерны множественные очаги симпластообразования в легких (рис. 2). Такого рода гигантоклеточная пневмония развивается на фоне иммунодефицитных состояний у детей в результате неконтролируемого размножения вируса и обычно фатальна.
Нередко возникает диарея — следствие вирусиндуцированного поражения энтероцитов. Но опаснее всего поражение центральной нервной системы. По данным электроэнцефалографии вовлечение ЦНС в инфекционный процесс наблюдается у половины детей, но, как правило, заканчивается благополучно. Лишь в одном из 1000—2000 случаев развивается острый коревой энцефалит. Он смертелен для 5—30% детей; у 20—40% выживших остаются тяжелые последствия (глухота, умственная отсталость, приступы судорог). Еще более драматичным является то, что энцефалит возникает на фоне выздоровления (чаще через 6 дней после начала сыпи) и не связан с тяжестью коревого синдрома. Инфекционный вирус редко выделяется из головного мозга, что позволяет думать об иммунологически опосредованной патологии, подобно поражению кожи и слизистых оболочек.
Неврологические осложнения этим не исчерпываются. Очень редким (1:300000—1:1000000), но символичным для вирусной персистенции заболеванием является подострый склерозирующий панэнцефалит (ПСПЭ). Он развивается через 5—15 лет после коревой инфекции, прогрессирует в течение 6—9 мес (иногда трех лет) и неизбежно ведет к смерти на фоне нарастающих психических и двигательных расстройств. В крови и спинномозговой жидкости определяются высокие титры противокоревых антител. В дегенерирующих нейронах можно найти коревые белки и РНК, а сам вирус удается выявить при сокультивировании с чувствительными клетками. По ряду признаков ПСПЭ-вирус отличается от классического вируса кори, являясь его дефектным вариантом с ослабленным репликативным потенциалом. Это связано с рядом мутаций, главная из которых вызывает дефект по М-белку, препятствующий созреванию вирионов.