Содержание
С поверхностными белками (H, N) связана высокая антигенная изменчивость вирусов гриппа. Она определяет рецидивы эпидемической агрессивности вируса типа А, меньше проявляется у вируса типа В и, по-видимому, не имеет практического значения для гриппа С (см. ниже).
Репликация
Вирус проникает в клетку путем эндоцитоза после связывания гемагглютинина с сиалированными клеточными рецепторами. Фагосома (эндосома), в которую заключен вирус, сливается с лизосомами. Кислая среда внутри фаголизосомы меняет конформацию гемагглютинина, обнажая пептиды, вызывающие слияние вирусной и эндосомальной мембран. Благодаря этому нуклеокапсид освобождается от суперкапсида и выходит в цитоплазму. Отсюда вместе с М-белком он быстро транспортируется в ядро, где уже через несколько минут после заражения появляются первые РНК-транскрипты. Число разновидностей мРНК соответствует количеству сегментов геномной РНК, хотя спектр синтезируемых белков несколько больше: к структурным белкам вириона добавляется 3 неструктурных белка, принимающих участие в репликации вируса. Расширение генетической емкости РНК достигается благодаря изящному механизму — повторной трансляции одной и той же мРНК со сдвигом рамки считывания ее генетического кода.
Ортомиксовирусы едва ли не единственные из РНК-вирусов, манипулирующие со своей геномной молекулой в ядре зараженной клетки. Клетки не умеют транскрибировать РНК, поэтому операции такого рода обязан производить сам вирус. Толчок дает полимеразный комплекс вириона, позже начинают работать вновь синтезированные ферменты. Белки РВ1 и РВ2 обеспечивают копирование геномных фрагментов с образованием неполных и полных по длине транскриптов. Укороченные копии выполняют функции мРНК. Для этого они подвергаются доработке, обретая облик, привычный для мРНК клетки (присоединение к 5`-концу шапочки из метилированных нуклеотидов и полиаденилирование 3`-конца). Не располагая собственными ресурсами для прямого решения этой задачи, вирус находит обходной путь. Он подключает свою эндонуклеазу (возможно, белок РА), которая откусывает недостающие фрагменты от мРНК клетки-хозяина и переносит их на вирусные РНК-транскрипты. Столь необычное и тонкое проявление молекулярного паразитизма объясняет природу внутриядерной репликации вирусов гриппа: только здесь они могут найти свежие (еще не связавшиеся с белком) мРНК, чувствительные к вирусной эндонуклеазе.
Полные по длине РНК-транскрипты не модифицируются и поэтому не могут работать как мРНК. Они остаются в ядре и служат матрицей для образования репликативных РНК, идущих на построение новых нуклеокапсидов. В этом принимает участие РНК-полимераза, образующаяся при помощи первых мРНК.
Готовые мРНК переправляются в цитоплазму, где транслируются на рибосомах в вирусные белки. Часть синтезированных белков вновь переходят в ядро и после комплексации с репликативными РНК образуют нуклеокапсиды. Гемагглютинин и нейраминидаза включаются в плазматическую мембрану, готовя фрагменты будущего суперкапсида. С внутренней стороны липидного бислоя к ним присоединяется М-белок, который ориентирует закрепление нуклеокапсидов в зонах почкования новых вирионов. При отделении от клеток вирус получает суперкапсид с его важнейшими атрибутами — гемагглютинином и нейраминидазой (рис. 4, 5).
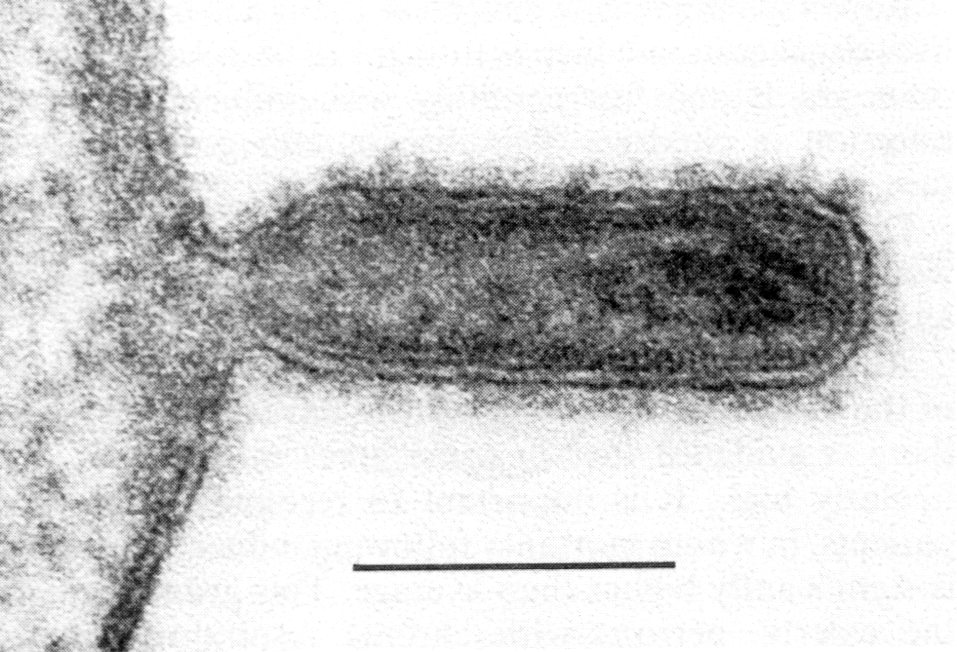
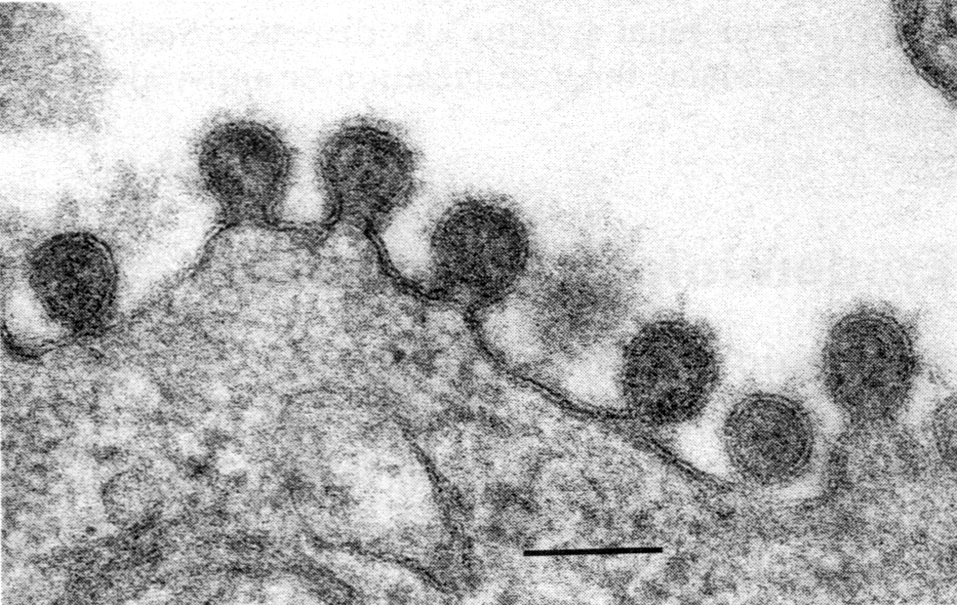
соответствует 100 нм
Как уже говорилось, гемагглютинин синтезируется в виде предшественника, который активируется клеточными протеазами (здесь корень формальной логики, породившей иллюзию о возможности лечения вирусных инфекций банальными ингибиторами протеаз). Сбрасывание созревших вирионов и их распространение на соседние клетки усиливается нейраминидазой. Репликация в чувствительных клетках происходит очень быстро, и уже через 6—8 ч в среде появляется вирусное потомство.
Экология
Кроме человека в естественных условиях вирусы гриппа инфицируют млекопитающих и птиц. Еще в 1931 г., т.е. за два года до открытия человеческого вируса, Р. Шоуп выделил пневмотропный вирус при массовых заболеваниях свиней. Тогда никто не предполагал, что это отпрыск возбудителя страшной «испанки», адаптировавшийся к новому хозяину во время пандемии 1918—1919 гг. Более того, оказалось, что вирус чумы домашних птиц (она протекает в виде септицемии, т.е. клинически совсем не похожа на грипп человека), обнаруженный в 1901 г., тоже близкий родственник вирусов гриппа человека. Эти открытия были оценены по достоинству с опозданием, когда появилась возможность серо- и генотипирования вирусных штаммов. Вирусы гриппа изолированы от лошадей, китов и тюленей. Обширным резервуаром служат птицы, выделяющие вирус с испражнениями.
Все изоляты от животных относятся к типу А. Они не патогенны для людей, но играют важную роль в эволюции эпидемических штаммов. Справедливо и обратное. Вирусы человека не вызывают эпизоотий, хотя пример с «испанским гриппом» свиней не исключает такой возможности. В эксперименте повышение вирулентности гриппозных штаммов наблюдается уже после нескольких пассажей на животных. Адаптация носит избирательный характер, распространяясь на узкий круг родственных хозяев.
Впрочем, в последние годы зарегистрированы случаи заражения людей птичьими штаммами вируса А. Реальная тревога возникла в 1997 г. в Гонконге, когда 18 человек напрямую заразились от домашних птиц вирусом H5N1; 6 человек погибло. В 2003 г. высоковирулентные штаммы птиц H5N1 и H7N7 спровоцировали фатальные случаи гриппа среди жителей Южной Азии. К счастью, они не получили распространения, но тем не менее очертили круг задач, решение которых необходимо для профилактики новых гриппозных пандемий.
Вирусы типов В и С являются сугубо человеческими и в естественных условиях не инфицируют других хозяев.
Патогенез
Вирусы гриппа проникают в респираторный тракт с каплями аэрозоля и частицами пыли. Чем мельче их величина, тем глубже проникает вирус; самые мелкие частицы достигают бронхиол и альвеол. Главным местом размножения вируса служат реснитчатые и бокаловидные клетки слизистой оболочки верхних дыхательных путей, но инфекция может охватить весь респираторный тракт — от носовых ходов до терминальных бронхиол и альвеол. Благодаря короткому репликативному циклу из каждой вирусной частицы к концу первых суток возникает огромное потомство — 1027. Это объясняет короткий инкубационный период (1—3 сут) и острейшее начало гриппа.
Повреждение зараженных эпителиоцитов индуцирует воспалительную реакцию, в которой преобладают макрофаги и лимфоциты. Подвергаясь активации, они секретируют множество флогогенных начал (прежде всего цитокинов), которые вызывают общую интоксикацию и поддерживают местный воспалительный процесс. В типичных случаях после внезапного недомогания, озноба с быстрым повышением температуры до 38—40оС, головной и часто мышечных болей развиваются симптомы поражения верхних (ринит, фарингит) и глубжележащих (ларингит, трахеобронхит) дыхательных путей. Общая интоксикация характерна для гриппа, но часто доминируют местные проявления. Наиболее тяжело протекает грипп А (хотя история эпидемий убеждает в вариабельности этого признака); вирусы типа В и, особенно, С менее агрессивны. В целом, опираясь только на клинику, отличить грипп от других ОРЗ практически невозможно. Без лабораторного подтверждения это удается лишь в сочетании с эпидемиологическим диагнозом, на фоне вспышки гриппа среди населения.