Содержание
Стратегия борьбы с гриппом очевидна — ее основой должна быть вакцинопрофилактика эпидемических разновидностей гриппозной инфекции. Для этого есть отличные технические возможности, которые базируются на знаниях о протективных антигенах вируса, эволюции вирусного генома, критериях антивирусной защищенности отдельных лиц и популяций. Неоднократно провозглашалось, что проблема почти решена, но результат всегда был скромнее претензий. Теперь понятно, что это не вина, а беда ученых. Мутанты и рекомбинанты вируса типа А, которые сочетают в себе антигенную новизну с высокой вирулентностью и не всегда своевременно отслеживаются эпидслужбами, создают массу трудностей, постоянно угрожая девальвацией вакцинных препаратов.
Слабость стратегии всегда рождает тактические ухищрения, часть из которых заведомо недобросовестны, а другие грешат формальной логикой и по сути являются тупиковыми. Немало подобных примеров знает и история гриппа. «Едва ли при какой-либо другой болезни было предложено и испробовано столько средств, сколько именно при инфлюэнции; но это обилие средств и доказывает, что нет ни одного верного». Об этом следует помнить, просматривая фармакологические справочники, где бесцеремонно рекламируются явно несостоятельные средства для лечения и профилактики гриппа. Что касается добросовестных заблуждений, то и они вовсе не безобидны, особенно с учетом экономической подоплеки.
Парамиксовирусы
- Вирион и структурные белки.
- Природа эпитопного консерватизма.
- Этапы репликации.
- Иммунитет и принципы этиотропной терапии.
- Вирусология и патология отдельных инфекций.
Собирательный термин миксовирусы (от греч. myxa — слизь) предложен Эндрюсом (C. Andrewes) в 1955 г. для вирусов гриппа, паротита и ньюкастлской болезни птиц. Он отражает сродство с муцинами и первоначально объединял оболочечные вирусы, способные связываться с мукополисахаридами и гликопротеинами, в том числе с сиалированными структурами клеток. Позже миксовирусы были разделены на два семейства — ортомиксовирусы (Orthomyxoviridae, вирусы гриппа) и парамиксовирусы (Paramyxoviridae), которые сильно отличаются друг от друга по биологии и строению вириона.
Парамиксовирусы человека включают вирусы парагриппа, паротита, кори и респираторно-синцитиальный вирус (РСВ). По современной классификации они входят в четыре родовых таксона:
- Paramyxovirus (вирусы парагриппа 1 и 3);
- Rubulavirus (вирус паротита, вирусы парагриппа 2 и 4);
- Morbillivirus (вирус кори)$
- Pneumovirus (респираторно-синцитиальный вирус).
Недавно появились сообщения о новом парамиксовирусе человека, который был изолирован (сначала в Нидерландах, затем в Австралии) от детей с острыми респираторными заболеваниями (ОРЗ). Он получил условное название «человеческий метапневмовирус» — hMPV (англ. human metapneumovirus). Полагают, что hMPV может быть причиной около трети ОРЗ с неустановленной этиологией.
К парамиксовирусам принадлежит также ряд вирусов млекопитающих и птиц (вирусы чумы собак и крупного рогатого скота, вирус болезни Ньюкастл птиц, возбудители ОРЗ домашних и диких животных). До недавних пор парамиксовирусы животных считались не опасными для человека. Но оказалось, что это не так. Появилось понятие зоонозные парамиксовирусы (род Henipavirus), с которыми связано несколько новых (англ. emergent) инфекций человека. Ошеломляющей явилась вспышка энцефалита в Малазии в 1998—1999 гг. Заболели 265 человек, 104 погибли. Причиной стал ранее не известный парамиксовирус, получивший название Nipah-вирус.
Заражение произошло от свиней, которые болеют Nipah-ОРЗ и гломерулонефритом, экскретируя вирус с респираторными секретами и мочой. Свиньи в свою очередь были инфицированы при контакте с одной из разновидностей летучих мышей, поддерживающих бессимптомную персистенцию Nipah-вируса. Для подавления вспышки пришлось уничтожить почти все поголовье свиней. Случаи заражения от больных людей не наблюдались.
Парамиксовирусы являются РНК-содержащими крупными оболочечными вирусами со спиральным типом симметрии. Это означает, что вирион (он имеет сферическую форму) имеет средний диаметр 150—300 нм с колебаниями от 100 до 800 нм и включает два главных компонента — нуклеокапсид, содежащий РНК-геном, и наружную липопротеиновую оболочку (суперкапсид) (рис. 1). Нуклеокапсид закручен в спираль, напоминающую винтовую лестницу, и обладает гибкостью, которая зависит от ионного состава и рН среды. Это позволяет ему скручиваться в компактную структуру (которая умещается в суперкапсид) и поддерживать плотный контакт с элементами РНК-полимеразного нуклеокапсидного комплекса.
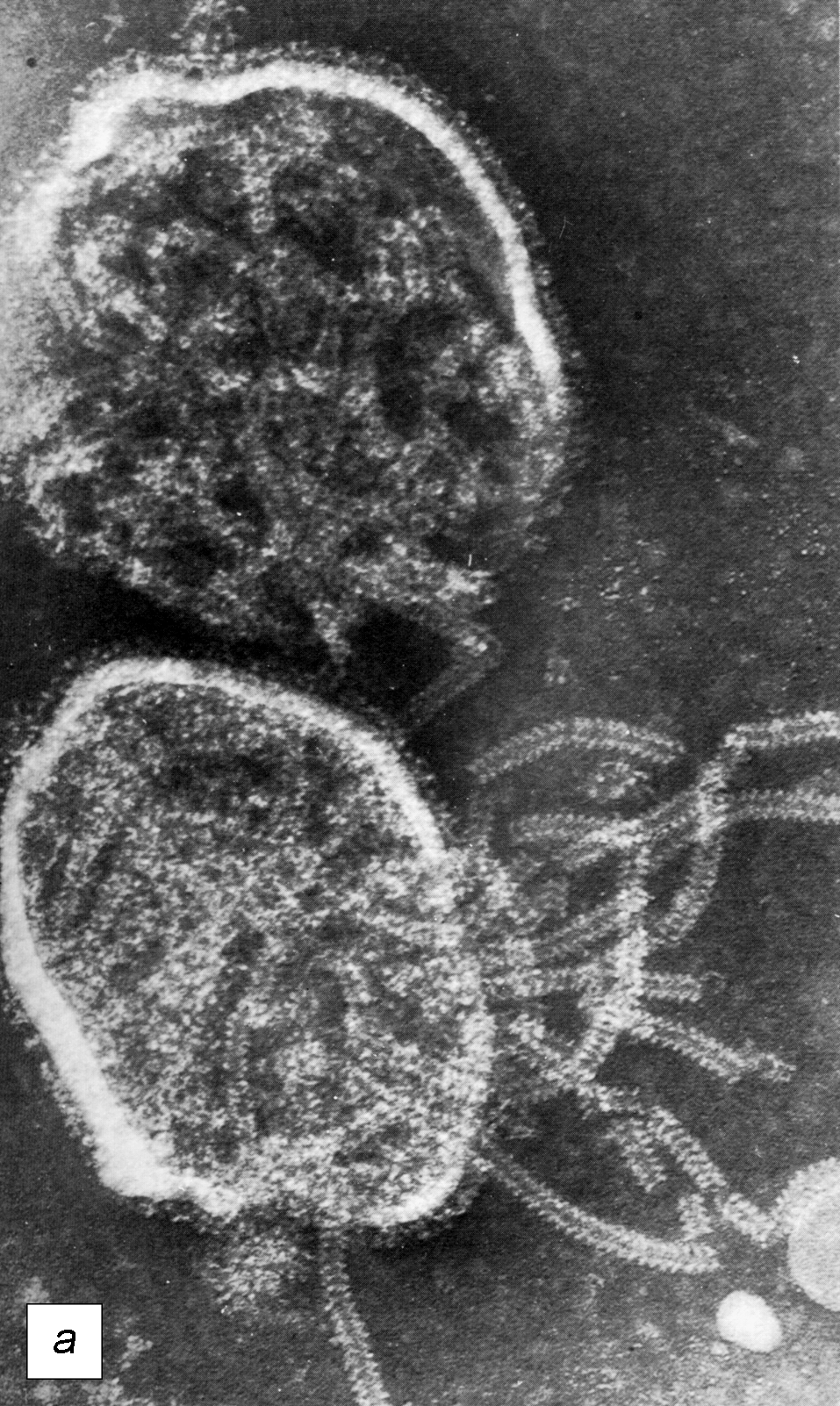
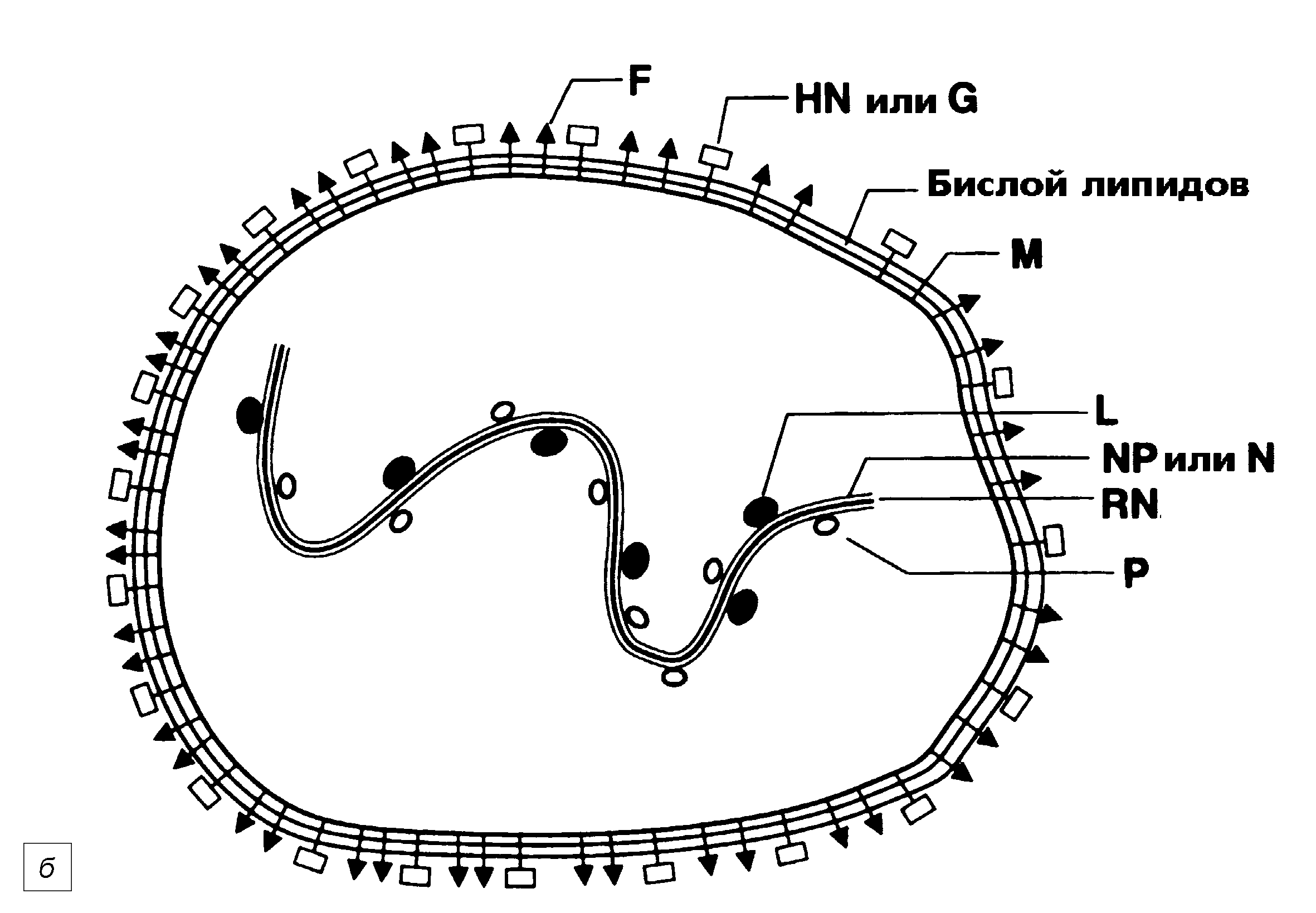
Рис. 1. Вирион парамиксовирусов: а — два вириона вируса парагриппа (электронограмма). На поверхности видна бахрома из суперкапсидных белков. Хорошо видна капсомерная (похожая на рыбий скелет) структура нуклеокапсида, частично выпавшего из нижнего вириона. Полоска равна 100 нм ; б — схема строения вириона парамиксовирусов. F — белок слияния, HN/H/G — рецепторный белок, M — матриксный белок, L и Р — белки РНК-полимеразногокомплекса, NP(N) — нуклеопротеин; RN — рибонуклеопротеин
На поверхности есть шипы, образованные двумя гликопротеинами. Более крупный наделен гемагглютинирующей и нейраминидазной активностью (вирусы парагриппа и паротита), только гемагглютинирующими свойствами (вирус кори) или лишен того и другого (РСВ). Значение этих различий не известно, но они относятся к числу классификационных признаков парамиксовирусов. Наличие или отсутствие гемагглютинирующей (Н) и нейраминидазной (N) активности отражены в акронимах — HN (вирусы парагриппа и паротита), Н (вирус кори) и G (от англ. glycoprotein; РСВ). В рецепции вирусов парагриппа и паротита участвуют сиаловые кислоты, для вируса кори главным клеточным рецептором служит CD46, на роль рецепторов для РСВ претендуют молекулы поверхностного глюкозаминоглюкана. Гемагглютинирующая и нейраминидазная активности не имеют отношения к рецепции: участки молекул, определяющие эти функции, структурно разобщены с сайтами связывания.
Второй гликопротеин, F (от англ. fusion — cлияние), формирующий более короткие шипы, вызывает слияние вирионной оболочки с мембраной клетки, обеспечивая прямое (минуя эндосому) поступление нуклеокапсида в цитоплазму. F-белок синтезируется в виде предшественника, F0, который сам по себе не активен, но обретает функцию слияния после ограниченного протеолиза. Его вызывают клеточные ферменты (фурины), относящиеся к семейству трипсиноподобных серинпротеаз. Протеолиз ведет к образованию двух субкомпонентов, F1 и F2, сохраняющих связь через дисульфидный мостик. Достаточно замены одного нуклеотида в f-гене, чтобы сделать F0 не чувствительным к протеолитической активации. Принципиально, что F0-расщепляющие ферменты в изобилии секретируются эпителиоцитами дыхательных путей. Это обеспечивает активацию F0 даже в тех случаях, когда он не успел подвергнуться внутриклеточному протеолизу. Функциональная сущность операции сводится к обнажению гидрофобного участка (пептида слияния) в составе F1, пенетрирующего клеточную мембрану.
Пептид слияния имеет структурное сходство не только у разных парамиксовирусов, но и гомологичен НА2-терминалу гемагглютинина (НА) вирусов гриппа, который выполняет такую же функцию. Но есть и принципиальное различие: F1 активен в условиях нейтральной среды и поэтому действует внеклеточно (на уровне плазматической мембраны); НА2 обретает активность после изменения конформации в кислой среде. Это происходит внутри эндосом после рецепторзависимого эндоцитоза вирионов. Только здесь (внутри клеток), внедряясь гидрофобным концом в эндосомальную мембрану, гриппозный гемагглютинин обеспечивает раздевание гриппозного нуклеокапсида и его выход в цитоплазму. Эти детали принципиальны для понимания взаимоотношений вирусов с клетками. Они служат ориентиром для разработки антивирусных препаратов, в частности структурных аналогов, блокирующих функцию пептида слияния. Сюда, казалось бы, вписываются и ингибиторы протеаз. Но сложность заключается в том, что в созревании вириона (F-белка) участвуют не вирусные ферменты, а протеазы клеток. Подавление их активности может быть небезразличным для хозяина.