Содержание
Большинство бета-гемолитических стрептококков человека являются болезнетворными. Они сравнительно редко выделяются от здоровых людей и всегда воспринимаются как потенциальная угроза для своего (обычно временного) хозяина и его окружения. Альфа-гемолитические стрептококки широко представлены в нормальной микрофлоре, особенно в ротовой полости (оральные стрептококки) и в толстом кишечнике (энтерококки; по современной классификации выделены в самостоятельный род Enterococcus). Это говорит о том, что альфа-гемолиз напрямую не связан с патогенностью, являясь скорее классификационным признаком, чем индикатором болезнетворности. Исключением является пневмококк (Str. pneumoniae) — один из важнейших патогенов человека. Он обладает мощной капсулой, которой лишены другие стрептококки, и именно она определяет его вирулентность. Негемолитические стрептококки входят в состав облигатной микрофлоры человека, но по своему видовому разнообразию уступают зеленящим стрептококкам. Они не играют роли в патологии человека или она минимальна.
Классификация стрептококков развивается в двух направлениях. Во-первых, было идентифицировано множество видов по биохимическим (ферментативным) признакам и факторам, претендующим на патогенность. Во-вторых, были разработаны методы серологической дифференциации стрептококков по групповым и типовым антигенам. Приоритет в этих исследованиях принадлежит американке Р.Ленсфилд, основополагающие работы которой (они относятся к тридцатым годам прошлого века) сыграли решающую роль в упорядочении знаний о стрептококках.
Деление на серологические (антигенные) группы за немногими исключениями основано на выявлении в реакции преципитации рамнозосодержащего полисахарида клеточной стенки, известного как С-субстанция. Преципитацией пользуются из-за того, что С-полисахарид часто экранирован другими компонентами клеточной стенки, и требуется экстракция, чтобы его обнаружить. В очищенном виде С-антиген является гаптеном, обретая иммуногенность в составе клеточных стенок. По его антигенным особенностям стрептококки разделены на 20 групп: A, B, C и т.д. Некоторые из них имеют статус вида, но есть и такие, которые включают несколько видов, дифференцируемых по другим признакам (например, группа С разделена на 4 вида). Принципиален факт: главный из медицинских стрептококков, Str. pyogenes, представляет единую серогруппу А, т.е. патогенетическая и серологическая классификации полностью совпадают. Известны стрептококки, которые лишены С-субстанции и классифицируются вне общей серологической схемы. К ним относится пневмококк и большинство зеленящих стрептококков, входящих в состав нормальной микрофлоры ротовой полости.
Бета-гемолитические стрептококки
Способностью лизировать эритроциты обладают стрептококки групп А, В (большинство штаммов), С, F и G. Это связано с продукцией гемолизинов, которые у стрептококка называются стрептолизинами. Дифференцируют два стрептолизина, S и О, которые различаются по чувствительности к кислороду: стрептолизин S (от англ. stable — устойчивый) не инактивируется в присутствии кислорода, стрептолизин О (от англ. oxygen-sensitive — чувствительный к кислороду) активен только в анаэробных условиях. Поэтому в обычных культурах (выращенных не в анаэростатах) гемолитическая активность обусловлена стрептолизином S. Впрочем, патогенетическая роль стрептолизина О, по-видимому, более значительна. Он обладает выраженной токсичностью для многих клеток (особенно для кардиомиоцитов), иммуногенен и антитела к нему (антистрептолизин О) служат индикатором активности стрептококкового процесса. Стрептолизин S менее токсичен и, не обладая иммуногенностью (отсутствие антител), не поддается специфической нейтрализации.
Большинство стрептококковых инфекций человека связаны с группой А, т.е. с Str. pyogenes, который в отличие от других бета-гемолитических стрептококков паразитирует только в организме человека. Гораздо реже сходные заболевания вызывают стрептококки В, С и G. Стрептококк группы В (Str. agalactiae) отличается своеобразной экологией — склонностью к паразитированию в родовом канале женщин; это делает его одним из важных факторов перинатальной патологии (см. ниже).
Str. pyogenes (группа А)
Антигены и факторы патогенности. Кроме С-группового полисахарида (группа А), в клеточной стенке Str. pyogenes содержатся еще несколько антигенов, по которым можно дифференцировать штаммы. Важнейшим из них является М-белок, структурно оформленный в виде филаментов, напоминающих пили/фимбрии грамотрицательных бактерий. По его антигенным особенностям идентифицировано более 80 М-типов (М-сероваров) пиогенного стрептококка, причем за редким исключением каждый из них содержит единственный вариант М-белка. М-белок (от англ. mucoid, так как колонии М-позитивных штаммов имеют мукоидную, т.е. слизистую консистенцию) представляет суперспирализованную молекулу, которая состоит из четырех участков (A—D), образующих филаментозные выросты, и двух фрагментов, которые заякоривают М-белок на мембране и в клеточной стенке (рис. 3). Эпитопная вариабельность связана с А-фрагментом, остальные участки более консервативны, что позволяет надеяться на создание универсальной противострептококковой вакцины.
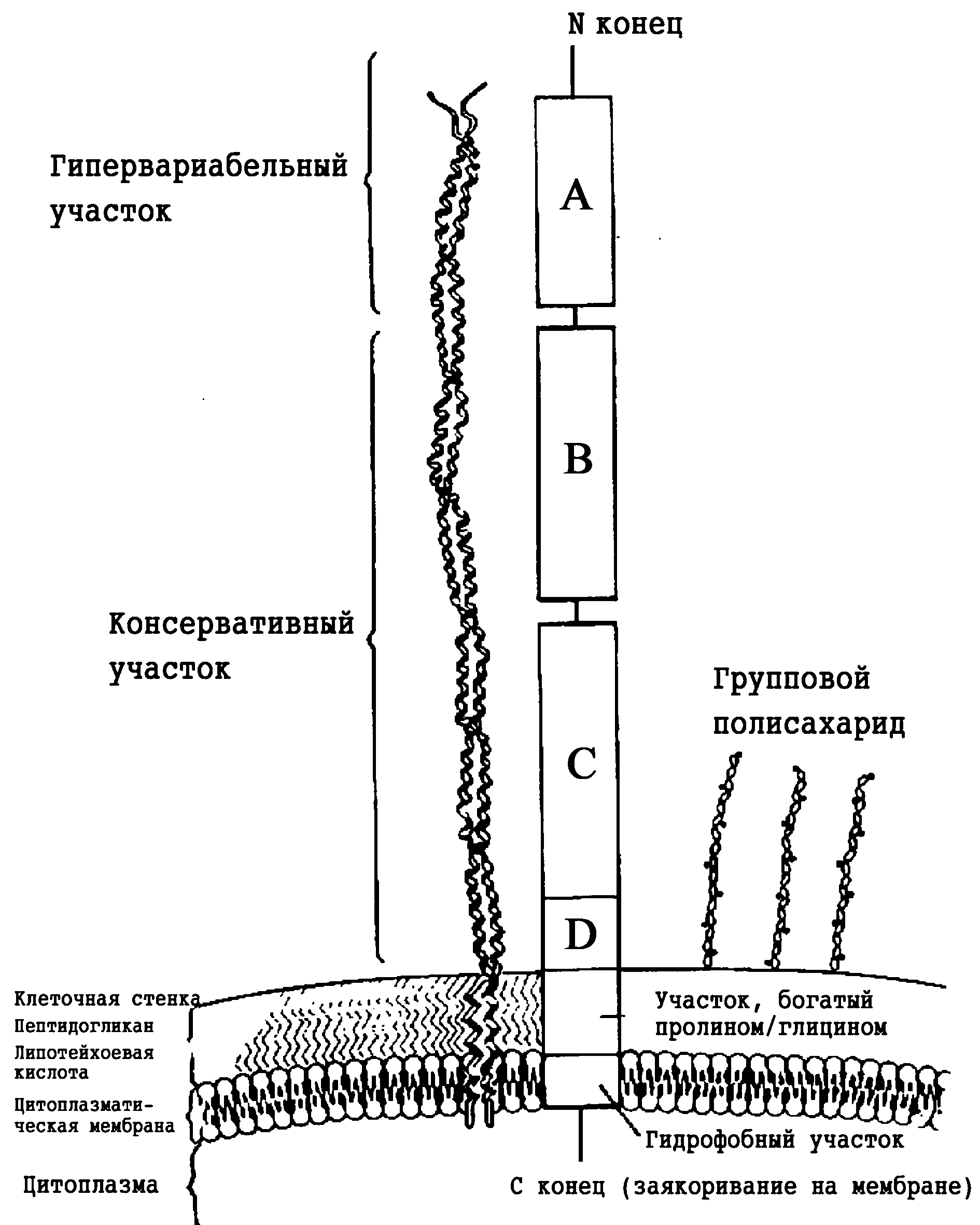
Типирование М-антигена оказалось полезным в эпидемиологических исследованиях, но главным явилось открытие его решающей роли в протективном иммунитете. Оказалось, что консервативные фрагменты М-антигена связывают фибриноген и Fc-участок IgG, формируя псевдокапсулу, экранирующую бактерии от фагоцитов. Антифагоцитарную активность М-белка дополняет фиксация сывороточного Н-фактора. Конкурируя с фактором В, он блокирует активацию альтернативного каскада комплемента, снижая эффективность антибактериальной реакции в опсонофагоцитарной системе. Снимая этот эффект, анти-М-антитела усиливают бактерицидность фагоцитов (нейтрофилов). М-белок имеет сайты связывания для фибронектина и CD46, способствуя адгезии стрептококка на эпителиальных клетках. Есть данные, что эта функция зависит от М-серотипа, отражая экологические варианты бактерий — респираторный и кожный стрептококки. Но и в этом случае не все ясно: некоторые высокоинвазивные и токсигенные (скарлатинозные) штаммы пиогенного стрептококка лишены М-подобных белков.
Узкая специфичность анти-М-антител определяет неудачную для человека иммунологию стрептококковых инфекций: иммунитет формируется раздельно для каждого из 80 М-сероваров Str. pyogenes. Кроме того, М-антигены обладают слабой иммуногенностью, и антитела к ним (по крайней мере с нейтрализующими свойствами) появляются с опозданием. Содержание М-белка снижается при культивировании бактерий на питательных средах, но быстро восстанавливается при пассировании на мышах. Это говорит о том, что стрептококки способны улавливать сигналы из внешней среды, контролируя образование фактора, играющего центральную роль в вирулентности. Установлено, что гены М-протеина (emm), входят в состав регулона, транскрипция которого находится под контролем позитивного регулятора mga, множественного генетического регулятора транскрипции стрептококка группы А. Экспрессия mga контролируется негативным регулятором Nra, который тормозит образование различных факторов патогенности стрептококка, в частности фибринонектинсвязывающего белка F2 и коллагенсвязывающего белка Сра. При ангине способность к продукции М-антигена падает по мере выздоровления, что лишний раз подчеркивает его отношение к агрессивности Str. pyogenes.
Кроме М-белка, к факторам антифагоцитарной защиты относятся стрептолизины, которые лизируют лейкоциты, подавляя фагоцитоз, и протеиназу, разрушающую С5а-фактор комплемента — мощный хемоаттрактант для нейтрофилов, способствующий их экстренной мобилизации в зону бактериальной инвазии. Ослабление этих механизмов снижает устойчивость к пиогенной инфекции, создавая предпосылки для опережающего размножения бактерий.
Кроме протеина М на поверхности многих штаммов имеются его рудименты — родственные и иммуноглобулинсвязывающие белки. Они кодируются теми же генами (emn или emn-related гены) и подобно М-белку связывают Fc-фрагмент IgG, имитируя защиту и, возможно, снижая опсоническую активность антител. Однако не все инвазивные и токсигенные штаммы располагают такими белками.
Большинство штаммов Str. pyogenes располагают капсулой, построенной из гиалуроновой кислоты. Так как гиалуроновая кислота входит в состав собственных тканей хозяина (это один из основных компонентов соединительной ткани), такая капсула лишена иммуногенности и не может быть атакована опсонинами. Теоретически это делает стрептококк неуязвимым, но он сам же лишает себя такого преимущества, продуцируя гиалуронидазу — фермент, разрушающий гиалуроновую кислоту. Поэтому патогенетическая (антифагоцитарная) роль капсулы сомнительна. Возможно, она способствует стабилизации инфекта в момент инвазии, но не имеет значения на последующих этапах инфекции. В частности, связываясь с гиалуронидатным рецептором CD44, капсула может быть одним из множества адгезинов, необходимых для закрепления стрептококка в верхних дыхательных путях.
Ненадежность собственной капсулы в известной мере компенсируется способностью Str. pyogenes фиксировать на своей поверхности белки хозяина (альфа-2-макроглобулин, фибриноген, плазминоген, фибронектин, витронектин, коллаген), создавая подобие капсулы.
В этой связи следует сказать и о липотейхоевой кислоте, пронизывающей клеточную стенку стрептококков. По инерции считают, что она играет едва ли не главную роль в закреплении (адгезии) бактерий на эпителии слизистых оболочек, связываясь с фибронектином, покрывающим эпителиоциты. Однако исследования с искусственными мутантами стрептококка заставляют усомниться в этом, выдвигая на первый план другие факторы адгезии (см. ниже).