Содержание
Существенную роль в выживании туберкулезной палочки играют сидерофоры — бактериальные факторы, конкурирующие с хозяином за железо. Для микобактерий это особенно важно, так как, будучи аэробами, они должны активно синтезировать железосодержащие ферменты. Макрофаги (наряду с гепатоцитами) являются главным резервуаром железа, избыток которого резервируется в виде ферритина. Содержание железа в альвеолярных макрофагах в 100 раз выше, чем в моноцитах крови, что, безусловно, содействует их колонизации микобактериями туберкулеза. Последние обладают уникальным тандемом сидерофоров — экзохелином и микобактином. Экзохелины (пептиды) извлекают ионы железа из микроокружения и передают микобактинам (липиды клеточной стенки), которые обеспечивают накопление и транспорт железа внутрь бактерий.
Таким образом, для выживания внутри макрофагов туберкулезная палочка использует сложную тактику, и только совокупность различных приемов обеспечивает ее агрессивность. Этим вирулентные штаммы отличаются от авирулентных, а также от других микобактерий. Повторяя туберкулезную палочку по ряду свойств, имеющих отношение к болезнетворности, они лишены их полного набора. Арсенал реально действующих начал не ясен и скорее всего раскрывается лишь в конфликте с хозяином. Не исключено, что некоторые факторы образуются только внутри зараженных клеток, как это происходит у других бактерий, предпочитающих внутриклеточный образ жизни. Например, сальмонеллы, паразитируя внутри макрофагов, дополнительно экспрессируют более 40 генов.
Гранулема. Взаимодействие между туберкулезной палочкой и макрофагами инициирует базисный для туберкулеза процесс — воспаление гранулематозного типа. Сама по себе гранулема не уникальна для туберкулеза. Сходные реакции возникают при других хронических инфекциях (бруцеллез, сифилис, кокцидиоидомикоз, лейшманиоз и пр.), а также при внедрении любых трудно метаболизируемых материалов. Например, гранулемы служат основой пневмоний, развивающихся при длительном вдыхании минеральной пыли. Множественные очаги гранулематозного воспаления образуются после внутривенных инъекций зимозана (полисахарид клеточной стенки дрожжей), угольных частиц и пр. Еще Кох обратил внимание на появление гранулем после введения животным убитых туберкулезных палочек или их дериватов. Такие реакции не перерастают в процесс, так как инъецируемый материал лишен способности к самовоспроизведению.
Зарождаясь тотчас после инфицирования, туберкулезные гранулемы получают мощный импульс с появлением Т-лимфоцитов, сенсибилизированных к возбудителю. Неспецифическая (доиммунная) гранулема трансформируется в специфическую (постиммунную), обретая признаки, характерные для туберкулеза. Именно с этого момента гранулема называется туберкулом. Подчиняясь диалектике воспаления, она служит не только рычагом саногенеза, но и механизмом повреждения. Следовательно, чтобы понять туберкулезный процесс, необходимо знать факторы, побуждающие гранулему к патогенетически значимой эволюции.
Неспецифическая (доиммунная) гранулема. Первичное заражение макрофагов не проходит бесследно. Добившись права на внутриклеточную репликацию, туберкулезная палочка вызывает их повреждение — пусть незначительное, но достаточное, чтобы стать центром для притяжения моноцитов крови, которые, попав в зону инфекции, трансформируются в макрофаги. Главным инструментом первичного повреждения служат липиды клеточной стенки с мембранотоксическим эффектом. Аппарат пиогенного (нейтрофилзависимого) воспаления после краткой вспышки практически бездействует. На протяжении всего процесса нейтрофилы остаются беспомощными свидетелями развертывающихся событий, располагаясь в небольшом количестве по периферии гранулемы. Если они и проявляют активность, то это скорее приносит вред, способствуя протеолизу некротических масс, который выгоден возбудителю.
Объяснение макрофагальной доминанты следует искать в природе микобактерий. Проникая в клетки, они исчезают из поля зрения эффекторов острого воспаления (комплемента, нейтрофилов, а затем и антител), которое гаснет, не получая стимулов к развитию. Медленное размножение туберкулезных бактерий и отсутствие у них ярких деструктивных начал снижает вероятность острого конфликта и в дальнейшем.
Стимулом к воспалению служит не только ортодоксальное повреждение клеток. Компоненты клеточной стенки (микозиды, сульфатиды, корд-фактор, липоарабиноманнан) побуждают макрофаги к секреции цитокинов и других флогогенных молекул, которые, привлекая моноциты, усиливают эффекторные ресурсы гранулемы и дирижируют событиями внутри нее. Кажется странным, но вирулентные штаммы в этом отношении менее активны. Например, их липоарабиноманнан не только не возбуждает, но даже блокирует секреторную реакцию макрофагов. Однако это формальное противоречие. Задержка воспалительного процесса способствует опережающему размножению бактерий, невольно содействуя стабилизации инфекта. Имеет значение и то, что туберкулезная палочка (тот же липоарабиноманнан), снижая чувствительность макрофагов к гамма-интерферону, ослабляет HLA-зависимое представление антигенов и пролиферацию Т-лимфоцитов.
Так или иначе, но доиммунная гранулема не останавливает инфекцию. Из легких туберкулезная палочка проникает в регионарные лимфатические узлы и далее в кровоток, получая возможность для диссеминации. Этому содействует сродство к макрофагам, присутствующим во всех тканях. За редким исключением дело ограничивается преходящими гранулематозными (в терминологии фтизиатров — параспецифическими) реакциями, повторяющими картину первичного очага инфекции. Сама по себе бактериемия на этом этапе не опасна, но она создает условия для персистенции возбудителя и его отсроченной реактивации в различных органах. С клинической точки зрения доиммунная фаза соответствует инкубационному периоду, так как никаких реальных симптомов здесь не возникает.
Специфическая (постиммунная) гранулема. Стратегическим итогом неспецифической гранулемы является развитие иммунного ответа против туберкулезной палочки. Реакция стартует в регионарных лимфоузлах, куда возбудитель и его антигены транспортируются (макрофагами, дендритными клетками) из первичной гранулемы. Дальнейшие события тесно связаны со специфическим воспалением, в основе которого лежит аллергическая реакция на микобактериальные антигены. Представления об аллергии как о необычной чувствительности к обычным (безвредным для нормального, т.е. несенсибилизированного организма) антигенам зародились именно при изучении микобактерий туберкулеза.
Все началось с наблюдений Коха. Он обнаружил, что у инфицированных животных чувствительность к туберкулезной палочке и ее продуктам гораздо выше, чем в контроле. Феномен Коха сводится к следующему. При подкожном заражении здоровой морской свинки культурой туберкулезной палочки видимые изменения на месте инъекции в первые дни отсутствуют. Лишь спустя 10—14 дней возникает плотный, изъязвляющийся узелок, который не заживает до гибели животного от генерализованного туберкулеза. Но если туберкулезные бактерии (живые, убитые) или их дериваты ввести внутрикожно или подкожно зараженному животному, то воспалительная реакция возникает уже через 24—48 ч. Ей сопутствуют общая интоксикация и обострение отдаленных очагов.
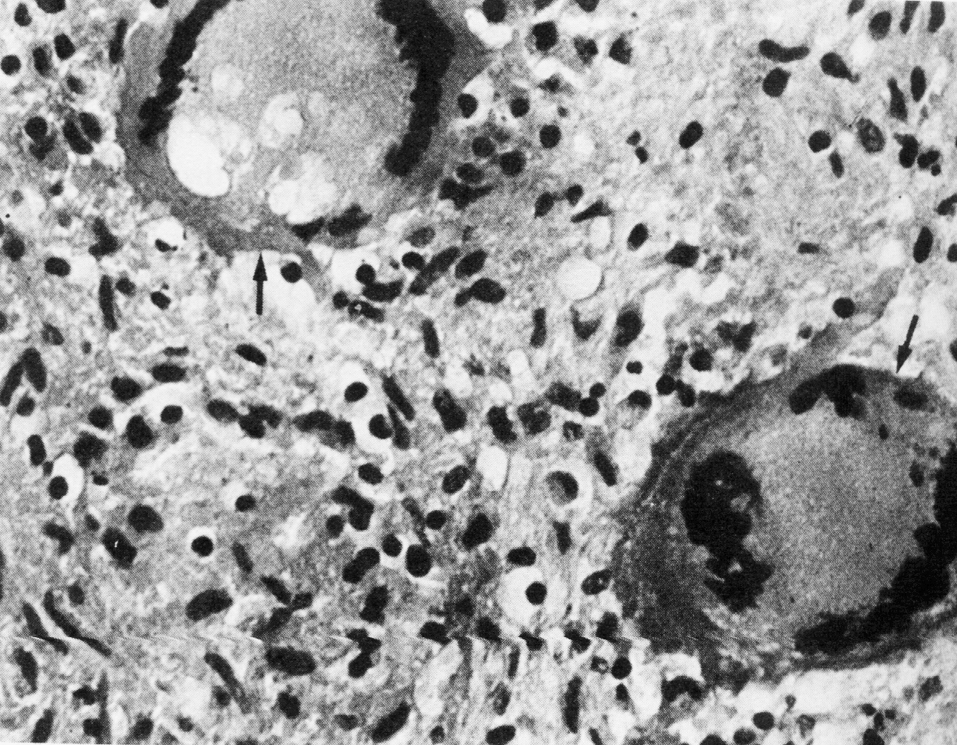
Отсроченный ответ при заражении здорового организма объясняется тем, что бактерии, которые размножаются в зоне инокуляции, до определенного момента сохраняют патогенетическую инертность. Ее нарушает сам хозяин, который после иммунной перестройки провоцирует гиперергическую (аллергическую) реакцию против возбудителя. В этом повинны сенсибилизированные Т-лимфоциты, круто меняющие ход событий. Они превращают доиммунную гранулему в мощный эффекторный аппарат, наделяя реакцию морфологией и агрессивностью истинного туберкула. Гранулема увеличивается в размерах за счет обильного притока новых клеток и объединения соседних бугорков. В ней появляются гигантские многоядерные клетки Лангерганса—Пирогова (результат слияния активированных макрофагов), макрофаги трансформируются в эпителиоидные клетки, центр гранулемы подвергается казеозному (творожистому) некрозу, по периферии накапливаются фибробласты, побуждаемые к секреции коллагена и отграничению зоны конфликта соединительнотканной (фиброзной) капсулой (рис. 4). Зрелый туберкул выглядит как плотный, сероватого (позднее желтоватого) цвета узелок диаметром 1—3 мм, не вылущиваемый из окружающей ткани.