Содержание
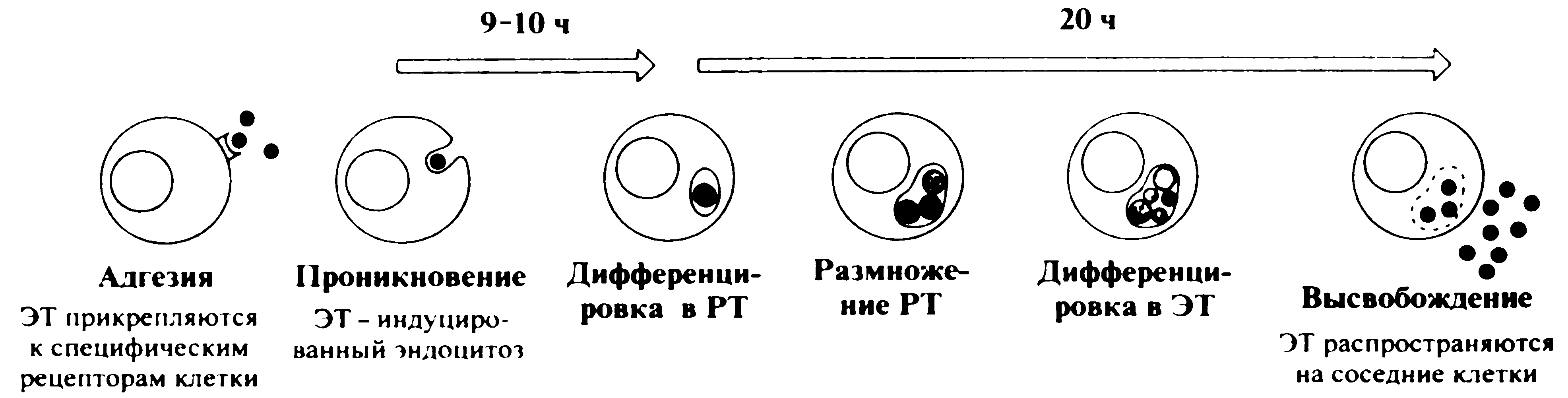
После поглощения ЭТ оказываются внутри фагосомы. Они вновь ведут себя нестандартно, блокируя слияние фагосомы с лизосомами, по крайней мере до окончания репликативного цикла. Благодаря этому хламидии избегают контакта с антимикробными факторами, получая возможность для беспрепятственного внутриклеточного развития. Оказавшись внутри клеток, ЭТ трансформируются в РТ. Это сопровождается структурной перестройкой мембран, которая повышает проницаемость клеток для АТФ и других питательных веществ. Говоря об уникальности жизненного цикла хламидий, следует помнить и о его известной аналогии со спорообразующими бактериями. С функциональной и экологической точек зрения ЭТ напоминают бактериальные эндоспоры. И те и другие предназначены для выживания во внешней среде, заражения новых хозяев и прорастания в вегетативные (т.е. способные к размножению) клетки. Для хламидий внешняя среда начинается тотчас после высвобождения из клеток. С этого момента стартует «звездный час» ЭТ с их относительной устойчивостью и способностью активно проникать в новые клетки.
Ретикулярные тела являются вегетативной формой хламидий. Отсутствие плотной оболочки делает их более хрупкими, чем ЭТ, но это и пробуждает в них жизнь, обеспечивая метаболические контакты с клеткой-хозяином. РТ многократно делятся, а затем (через промежуточные формы) превращаются в ЭТ. Это совершается не одновременно, а потому на протяжении всего цикла внутри клеток можно видеть РТ, ЭТ и промежуточные формы (см. рис. 2). При множественном заражении образуется несколько фагосом — вакуолей, содержащих ЭТ. В дальнейшем они сливаются вместе (Сh. trachomatis; см. рис. 1) либо дают начало дискретным внутриклеточным включениям (Ch. psittaci; рис. 3).
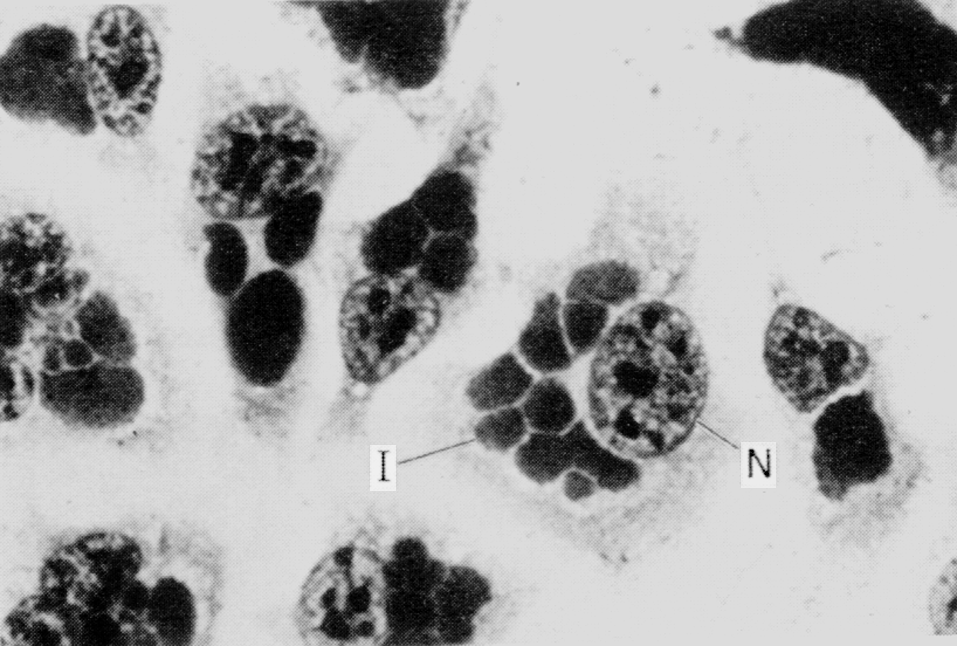
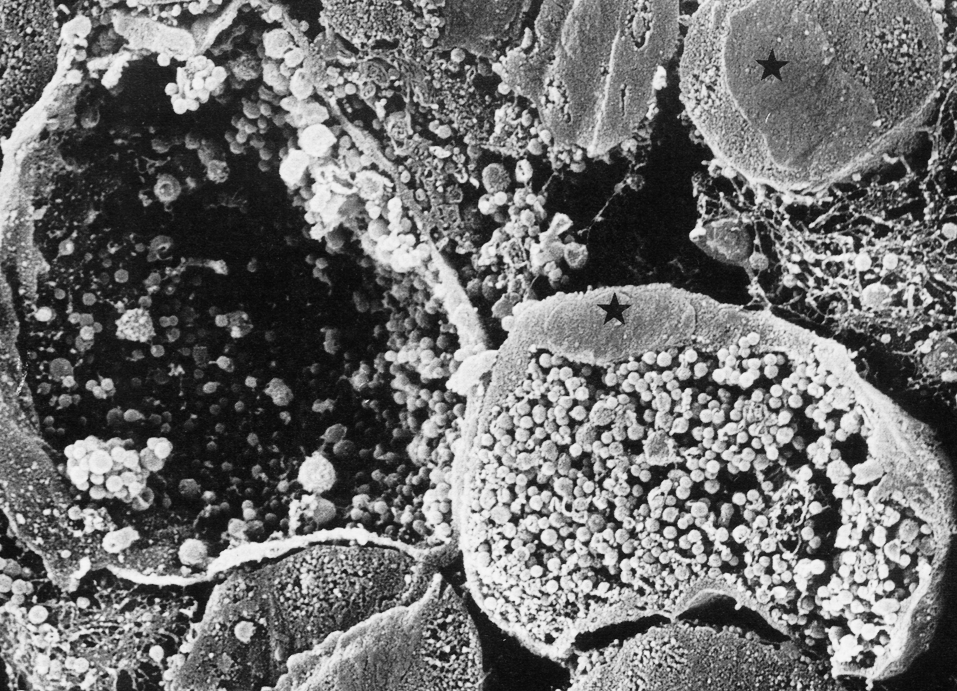
В конце цикла включения (по сути микроколонии хламидий) занимают большую часть клетки и содержат многочисленное потомство ЭТ (рис. 4). Растягивая стенку вакуоли, они разрывают ее, а затем и плазматическую мембрану. Цикл продолжается 40—48 ч, завершаясь гибелью клетки и выходом ЭТ на свободу (рис. 5).
Этому способствует и ослабление конструктивного метаболизма зараженных клеток, которые переключаются на обслуживание бурно реплицирующихся хламидий. В целом это логично объясняет деструктивный эффект хламидий, тем более что никаких специальных цитотоксинов у них не обнаружено. Не следует, впрочем, забывать о том, что при высокой инфицирующей дозе хламидии способны убивать клетки до репликации, т.е. перед тем, как возникает давление внутриклеточных включений, переполненных микробными частицами.
Репликативный цикл может быть искусственно растянут и даже приостановлен, если зараженные клетки поместить в неблагоприятные условия (например, лишить необходимых факторов питания) или добавить ингибиторы размножения хламидий (например, пенициллин). Хламидии не погибают, сохраняя способность к размножению после устранения негативных факторов. В известной мере это моделирует склонность хламидий к латентным (персистентным) инфекциям (см. ниже).
Классификация хламидий формально проста. Порядок Chlamydiales включает одно семейство (Chlamydiaсeae) c единственным родом (Chlamydia). Число видовых таксонов тоже весьма ограничено. До недавнего времени их было всего два — Ch. trachomatis и Ch. psittaci, в 1989 г. введен еще один вид — Ch. pneumoniae. О таксономической самостоятельности каждого из них говорит множество эколого-патогенетических, микробиологических и иммунохимических вариантов. Этому соответствует и слабое генетическое родство видовых таксонов хламидий — всего 5—10% гомологии ДНК.
В 1999 г. предложена новая классификация, привыкание к которой (если оно состоится) потребует времени. Классификация основана на генетических критериях, использующих степень гомологии в нуклеотидной последовательности генов 16S и 23S рибосомальных РНК: >95% для всех представителей одного рода, >90% — семейства и >80% — порядка/класса микроорганизмов. В соответствии с этим семейство Chlamydiaceae разделено на два рода: Chlamydia и Chlamydophila. В состав рода Chlamydia кроме Chlamydia trachomatis вошли Chlamydia muridarum и Chlamydia suis (патогены животных). Род Сhlamydophila составляют Chlamydophila pneumoniae (прежнее название Chlamydia pneumoniae) и варианты прежнего вида Chlamydia psittaci (Chlamydophila psittaci, Chlamydophila abortus, Chlamydophila felis, Chlamydophila pecorum и Chlamydophila caviae). Chlamydophila psittaci включает штаммы 8 сероваров, для которых основными хозяевами являются птицы. Все они могут передаваться человеку, вызывая пситтакоз (см. ниже).
Chlamydia trachomatis
Взаимоотношения с человеком, единственным естественным хозяином Ch. trachomatis (исключение составляют биовары Ch. muridarum и Ch. suis, вызывающие пневмонию у мышей и свиней), определяются способностью данного вида персистировать в цилиндрическом эпителии генитального тракта и конъюнктивы. Часто это протекает бессимптомно, однако в ряде случаев слабое, но рецидивирующее повреждение эпителия возбуждает и поддерживает воспалительную реакцию слизистых оболочек — основу хронической патологии, типичной для Ch. trachomatis.
Принято выделять три разновидности/биогруппы Ch. trachomatis: глазные, генитальные и хламидии венерической лимфогранулемы (ВЛГ). Последние обладают повышенной вирулентностью и способны выходить за пределы слизистых оболочек, поражая регионарные лимфатические узлы. Глазные и генитальные хламидии являются строгими эпителиальными паразитами, лишенными способности инвазировать глубжележащие ткани.
Эколого-патогенетическая неоднородность Ch. trachomatis обрела реальную основу в 1970-х гг., когда при помощи микроиммунофлюоресценции удалось серотипировать Ch. trachomatis. В настоящее время известно 18 cероваров, принадлежность к которым удивительным образом соответствует патогенетическим потенциям хламидий. Серовары А, В, Ва, С включают глазные хламидии, вызывающие трахому; D—K-хламидии служат причиной урогенитального хламидиоза и его экстрагенитальных осложнений; L-серовары (L1, L2, L3) являются возбудителями ВЛГ.
Трахома — это хронический конъюнктивит, в основе которого лежит гранулематозное воспаление конъюнктивы, захватывающее роговицу (кератоконъюнктивит). Начавшись в раннем детстве, заболевание прогрессирует на протяжении многих лет и полностью купируется лишь после зарастания очагов инфекции рубцовой соединительной тканью. Это тот случай, когда микробиологическое выздоровление дается ценой тяжелых структурных изменений, ведущих к потере функции, в данном случае зрения. Эпителий конъюнктивы человека — единственный природный резервуар возбудителя трахомы, его входные ворота и зона патогенетически значимого размножения. Трагический парадокс заключается в том, что сугубо местный процесс, который на первых порах доставляет мало беспокойств и вполне излечим, обретает катастрофическую необратимость. Впрочем, как и при других хламидиозах, процесс нередко ограничивается начальными проявлениями, не достигая тяжких последствий. Более того, возможны бессимптомное инфицирование и даже длительная персистенция без клинических проявлений.
Название болезни пришло из глубокой древности. В переводе с греческого «trachys» означает «шероховатый, неровный». Это связано с тем, что поверхность конъюнктивы выглядит бугристой из-за лимфоидных фолликулов (трахоматозных гранулем), которые формируются в субэпителиальной ткани. В запущенных случаях вся конъюнктива сплошь усеяна зернышками, тесно прилегающими друг к другу, так что получается картина вареного саго. Отсюда об активной фазе болезни говорят как о фолликулярном конъюнктивите, подчеркивая патогенетическую уникальность трахомы в сравнении с другими конъюнктивитами (кстати, одно из старых названий трахомы — зернистое воспаление глаз).