Содержание
Но мы забежали вперед — всему этому предшествует проникновение вириона в клетку, высвобождение (раздевание) вирионной ДНК и ее подготовка к транскрипции и репликации. Вирус прикрепляется к клеткам концами своих фибрилл. Природа клеточных рецепторов точно не установлена, но скорее всего это комплекс из нескольких белков, обеспечивающих мультивалентное связывание вириона с плазматической мембраной. В реакцию слияния вступают белки основания пентона, которые взаимодействуют с интегринами клеточной мембраны, приводя к эндоцитозу вирусрецепторного комплекса. В кислой среде эндосом вирион частично теряет свою капсидную оболочку и после выхода в цитоплазму при содействии микротрубочек проникает через поры ядерной мембраны в ядро, сбрасывая остаток гексонов и полностью освобождая ДНК от белков. Весь процесс раздевания вирусной ДНК занимает 1—2 ч.
Белки сердцевины замещаются клеточными гистонами с образованием хроматиноподобного комплекса, готового к экспрессии ранних вирусных генов. Среди последних есть лидер, Е1а, который транскрибируется первым и поэтому называется сверхранним. Один из трех кодируемых им белков выполняет функцию транскрипционного активатора, индуцируя транскрипцию для остальных ранних генов. Лидерству Е1а способствует его соседство с терминальным белком на конце ДНК, который служит затравкой для РНК-полимеразы. В репликации аденовирусов немало и других тонкостей, которые отражают сложные взаимоотношения не только с зараженными клетками, но и с организмом хозяина в целом. Отметим, в частности, способность аденовирусов нейтрализовать механизмы антивирусного иммунитета. Благодаря двухспиральным молекулам РНК они блокируют интерферонзависимую инактивацию фактора элонгации 2, участвующего в синтезе вирусных белков. Антиинтерфероновой активностью обладает и белок Еа1, блокирующий транскрипцию генов, индуцируемых интерфероном. Производные гена Е3 подавляют транспорт молекул главного комплекса гистосовместимости класса I (HLA-I) на поверхность зараженных клеток, тем самым защищая их от иммунного цитолиза.
Сборка вирионов происходит в ядре, куда структурные белки транспортируются из цитоплазмы после их синтеза на рибосомах и частичного объединения в капсомеры, т.е. в мультипептидные комплексы. Каждый гексоновый капсомер собирается из трех идентичных белков, пентон образуется из пяти одинаковых белков пентонового основания и трех белков фибриллы. В агрегации гексоновых субкомпонентов участвует формообразующий поздний белок (продукт гена L4), который не включается в состав вириона.
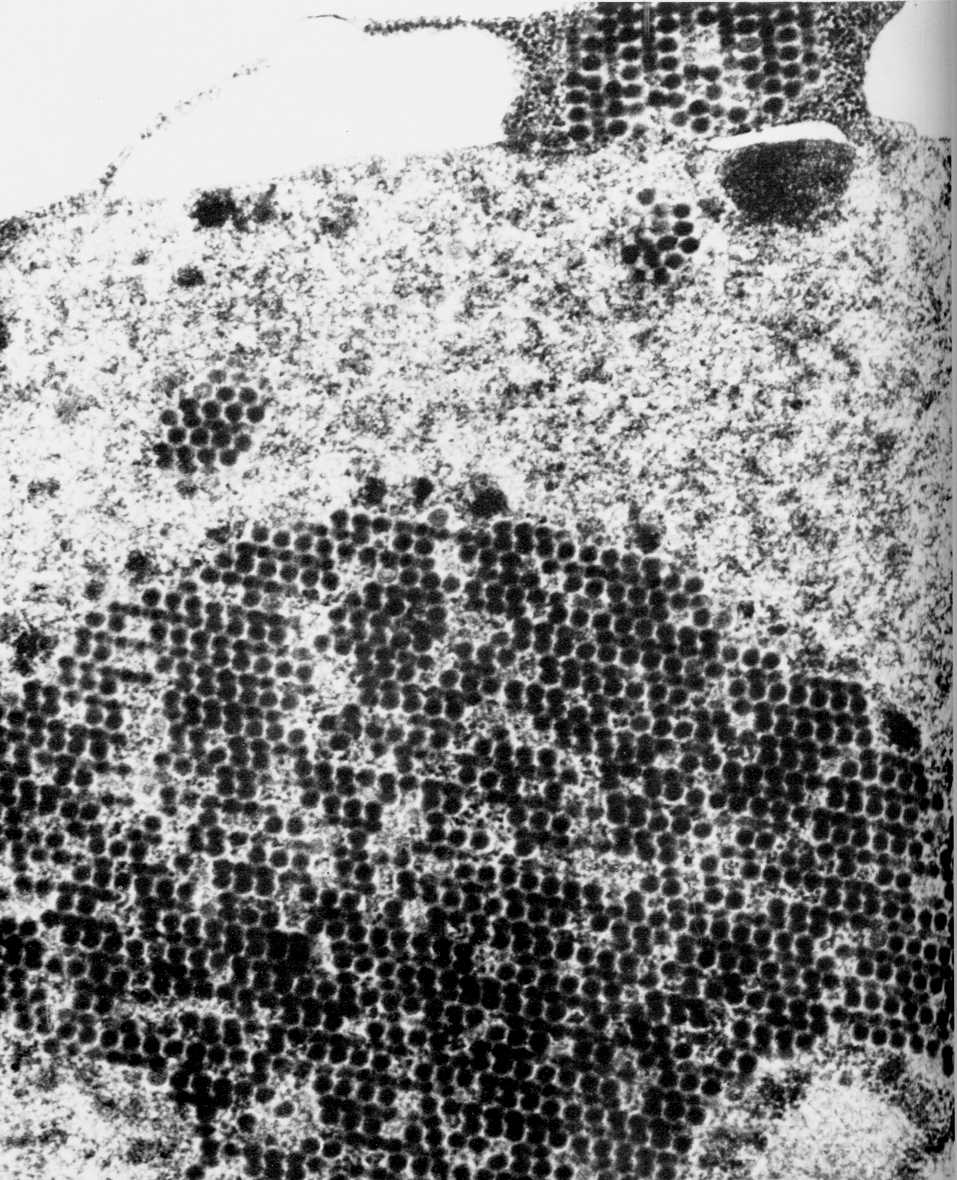
Оказавшись в ядре, капсомеры самоорганизуются в капсиды, куда включается вирусная ДНК. Конструкцию вириона завершает вирусная цистеин-протеиназа (продукт гена L3), которая доводит до финальной кондиции структурные белки. Общая продолжительность репликативного цикла составляет примерно 24 ч, но, как уже говорилось, новые вирионы не торопятся с выходом из клетки, дожидаясь ее гибели. Сборка вирионов происходит неэкономно. Структурные белки синтезируются в большом избытке, оставаясь неиспользованными. Они накапливаются в ядре в виде кристаллов вместе со скоплением зрелых вирионов (рис. 2).
Даже такое краткое описание аденовирусной инфекции демонстрирует ряд вирусспецифических факторов и механизмов, подавление которых могло бы приостановить развитие инфекционного процесса. Однако до сего времени ни один из них не использован в качестве мишени для этиотропной терапии.
Онкогенность
Несомненно, самым интригующим фактом является способность аденовирусов оказывать туморогенное действие на клетки. Принципиально, что за исключением экспрессии нескольких ранних генов этот эффект не имеет ничего общего с продуктивной инфекцией, т.е. с воспроизводством инфекционного вируса. Более того, туморогенность наблюдается только в непермиссивных или полупермиссивных системах, т.е. по сути отражает абортивную инфекцию, при которой не происходит полной экспрессии вирусного генома, а репликативные события ограничиваются синтезом ранних белков. В таких случаях клетки не погибают, а становятся бессмертными (иммортализуются), обретая злокачественные потенции, губительные для хозяина. К сожалению (или к счастью), прагматичный финал интриги разочаровал. Аденовирусные ДНК и мРНК не удалось найти ни в одной из опухолей человека. Ничего особенного не обнаружено и иммунологами: антитела к туморассоциированным антигенам (Т-антигенам), которые являются обязательным спутником опухолевого процесса, провоцируемого аденовирусами в опытах на животных, отсутствуют у больных злокачественными новообразованиями. Это означает, что роль аденовирусов в возникновении человеческих опухолей должна быть отвергнута. Онкогенность проявляется только в модельных опытах на животных и для ее реализации требуются достаточно искусственные условия. Тем не менее, изучение аденовирусов сыграло большую роль в становлении современных концепций, а аденовирусная инфекция остается одной из главных моделей молекулярной онкологии.
Прежде всего следует отметить, что все аденовирусы человека вызывают стабильные морфофункциональные изменения в культурах непермиссивных и полупермиссивных клеток грызунов, напоминающие то, что происходит при естественном канцерогенезе. Онкологи называют это трансформацией. Трансформированные клетки размножаются быстрее нормальных, менее требовательны к питательной среде, утрачивают «чувство локтя» (т.е. выходят из-под контроля контактной ингибиции) и поэтому растут хаотично, многослойно, а главное — обретают фенотип бессмертия, т.е. способность к бесконечным инвитровым пассажам, что может быть связано с инактивацией теломеразы — фермента, укорачивающего концевые отрезки ДНК при каждом делении клеток. Центральным событием является интеграция вирусной ДНК с хозяйским геномом. Для этого требуется лишь небольшой участок (8—14%) аденовирусной ДНК, расположенный на одном из ее концов. Здесь локализованы гены Е1а и Е1b, и именно их продукты (Т-антигены) ответственны за трансформирующий эффект. Этим объясняется то, что трансформацию можно вызвать не только цельным вирусом, но и его ДНК и ее фрагментами (большинство клеток содержит не более 20% аденовирусного генома). Т-антигены содержатся в основном внутри ядра, определяя особенности трансформированных аденовирусами клеток, по которым их можно отличить от опухолевых клеток иного происхождения.
Механизм трансформации не совсем ясен. Сравнение клеточных линий показало, что вставки вирусных ДНК-фрагментов в хромосомы носят случайный характер, т.е. лишены сайтов для предпочтительной интеграции. С этой точки зрения трансформацию трудно представить как результат инсерционного мутагенеза, т.е. как дестабилизацию клеточного генома вирусными вставками. Против этого говорит отсутствие сайтовой специфичности интеграционного процесса, с чем можно было бы связать извращение функции клеточных генов (онкогенов), контролирующих пролиферацию. Скорее всего эффект опосредован через взаимодействие аденовирусных белков с регуляторами роста и деления клеток. Вряд ли случайно, что среди продуктов трансформирующей области аденовирусного генома есть факторы, влияющие на апоптоз, т.е. на генетически запрограммированную гибель клеток.
Второй аспект проблемы связан с онкогенностью in vivo. Оказалось, что некоторые аденовирусы человека вызывают опухоли у грызунов (наиболее чувствительны новорожденные хомячки). По данному признаку аденовирусы делятся на высокоонкогенные, слабоонкогенные и неонкогенные (см. табл. 1). Вряд ли можно сомневаться, что в основе онкогенности лежит эффект трансформации клеток, создающей прецедент для бесконтрольного размножения. Об этом говорит развитие опухолей после введения животным «адекватно» трансформированных клеток. Вместе с тем критерии такой «адекватности» отсутствуют, и далеко не все линии трансформированных клеток обладают туморогенностью. В частности, не выявлено очевидной корреляции между какими-либо признаками трансформированных клеток и их онкогенностью для новорожденных хомячков. Трансформацию вызывают даже неонкогенные серотипы, но опухоли надежно воспроизводятся лишь с представителями подгрупп А и В. Несомненно, что онкогенный потенциал зависит от дополнительных изменений фенотипа, и не исключено, что, несмотря на формальное сходство, трансформация, вызванная разными вирусами, несет в себе неодинаковую агрессивность.