
2006 год
Если лечиться, будешь здоров через неделю, если нет — через 7 дней.
Рино- и коронавирусы
- Экология и патогенетический потенциал.
- Риновирусы:
классификация и многоликость;
репликация;
молекулярный и клеточный патогенез. - Коронавирусы: реальность и перспективы;
необычность репликации;
причины отсутствия прочного иммунитета.
Рино- и коронавирусы относятся к ОРВИ и специализируются на поражении верхних отделов дыхательного тракта. Одной из причин этого может быть их наибольшая активность при пониженной температуре (33—34оС), которая характерна для полости носа. Доминируя (как и другие респираторные вирусы) в раннем детстве, рино- и коронавирусы остаются главной причиной малой респираторной патологии, или простудных заболеваний (англ. cold) на протяжении всей жизни. Их символ — острый ринит (заразный насморк), к которому обычно присоединяется поражение носоглотки (фарингит), реже — параназальных синусов, среднего уха, трахеи. Признаки общей интоксикации (температура, слабость, озноб и пр.) отсутствуют или выражены слабо
Все это хорошо известно каждому по собственному опыту. Взрослые в среднем переносят не менее одной простуды в год, для детей (особенно посещающих детские дошкольные учреждения) показатель еще выше. Повышение чувствительности к ОРВИ-вирусам при различного рода переохлаждениях зафиксировано многовековой эмпирикой, однако до сих пор никто серьезно не пытался проанализировать причины этого явления. Можно, например, задуматься о реактивной гиперэкспрессии вирусных рецепторов на чувствительных клетках или о снижении количества сывороточных антител, мобилизуемых на поверхность слизистых оболочек из микроциркуляторного русла. Рассуждая о рино- и коронавирусах, следует учитывать и их психрофильность. Впрочем, значение простудного фактора, возможно, преувеличивается. По крайней мере, в контролируемых опытах искусственное переохлаждение не повышало чувствительности к риновирусам.
В США подсчитано, что из-за простудных заболеваний в год теряется 161 млн. дней нормальной трудоспособности, они вызывают 26 млн. дней пропущенных школьных занятий, 23 млн. дней освобождения от работы и 27 млн. обращений за врачебной помощью. Ежегодные затраты на лечение больных превышают 2 млрд. долларов. Рекомендовано более 800 лекарств «от простуды», которые, однако, не отменили классической формулы: если будешь лечиться, здоров через неделю; если нет — через 7 дней.
Риновирусы
Род Rhinovirus относится к пикорнавирусам (Picornaviridae). В названии семейства отражены два признака: минимальный (около 30 нм) размер вириона (лат. picus — миниатюрный, мелкий) и тип геномной молекулы — РНК (ribonucleic acid, RNA). Вирион пикорнавирусов лишен липидной оболочки (суперкапсида), т.е. является голым нуклеокапсидом. Капсид построен по типу кубической (икосаэдральной) симметрии из одинаковых блоков (капсомеры, или протомеры), в состав которых входит по четыре белка — VP1, VP2, VP3 и VP4 (от англ. viral proteins). РНК имеет позитивную полярность, т.е. ведет себя как мРНК, напрямую обеспечивая синтез вирусных белков.
Род Rhinovirus объединяет множество серотипов (иммунотипов), дифференцируемых по реакции нейтрализации в клеточных культурах. С учетом двух антигенных вариантов типа 1 (1А,В) и Hank-риновируса (Н), которому не присвоен официальный порядковый номер, число серотипов составляет 102. Все они так или иначе задействованы в респираторной патологии. Риновирусы вызывают 30—50% острых респираторных заболеваний, являясь абсолютным лидером среди возбудителей простуды — ринита (с этим связано родовое название — Rhinovirus) и фарингита. Естественным резервуаром является носоглотка и полость носа человека, прежде всего детей. Чувствительность к риновирусам достаточно высока, поэтому большинство случаев заражения имеет клинические последствия. К тому же для инициации процесса требуется ничтожное количество вируса.
Согласно эпидемиологическим наблюдениям и опытам на добровольцах, соотношение между клинически значимыми и бессимптомными риноинфекциями составляет 3:1. После острого пика (примерно 48 ч после заражения) выделение вируса быстро падает, продолжаясь не более 3 нед. В силу этих причин носительство риновирусов сильно лимитировано: они выделяются лишь от 2% здоровых людей.
Заражение скорее всего происходит контактным, а не аэрозольным путем. Риновирусы не удается обнаружить в воздухе помещений, где находятся инфицированные люди, а их содержание в респираторных секретах при кашле и чихании невелико. Это можно понять, если учесть, что аэрозоли, возникающие при разговоре, кашле и чихании, происходят главным образом из слюны (где концентрация риновирусов мала), а не из назальных секретов, содержащих максимальные титры вируса. В острой фазе риновирусы постоянно присутствуют на пальцах больных, откуда путем опосредованных контактов (рукопожатие, зараженные предметы) могут попадать на слизистую оболочку носа или конъюнктиву (и далее, через слезный проток, в носовую полость) других людей. Известно, что простудные заболевания вяло распространяются в коллективах, где общее воздушное пространство сочетается с ограниченностью контактов. Напротив, заражение легко происходит в семьях, когда тесные контакты неизбежны. Инфекцию часто приносят дети, посещающие организованные коллективы (ясли, детские сады).
Большинство риновирусов связываются с эпителиоцитами через хорошо известный рецептор ICAM-1 (от англ. Intracellular Adhesion Molecule). Экспрессия ICAM-1 усиливается медиаторами воспаления, что должно способствовать заражению клеток. Это может быть одной из причин неодинаковой чувствительности к риновирусам и ее периодических колебаний. Репликация происходит так же, как у других пикорнавирусов. Рецепция вириона возбуждает конформационные изменения капсидных белков, которые ведут к высвобождению вирионной РНК и ее трансмембранному переходу в клеточную цитоплазму. Здесь она тотчас подвергается трансляции, обеспечивая синтез полипротеина — предшественника всех (чуть более десятка) структурных/вирионных и неструктурных вирусных белков. Они образуются в результате поэтапного расщепления полипротеина двумя вирусными протеазами, которые вычленяются протеазой клетки или являются результатом аутокатализа. Все эти события, как и сборка новых вирионов, происходят в цитоплазме. Вирионы выделяются при цитолизе, который скорее всего возникает из-за подавления синтеза клеточных белков. Установлено, что риновирусные протеазы вызывают образование по крайней мере двух факторов, необходимых для транскрипции и трансляции клеточного генома (факторы IIIC и 4G).
Наиболее интенсивное размножение вируса происходит в задней части носоглотки. Первичной мишенью могут быть М-клетки — специализированные эпителиоциты, покрывающие аденоидные крипты. Они имеют максимальную плотность молекул ICAM-1, рецептирующих риновирусы. Возможно, конфликт зарождается именно здесь, но быстро распространяется на реснитчатые эпителиоциты носа. По времени это совпадает с первыми признаками заболевания — першение в горле, саднение в носу, чихание. Они появляются уже через 10—12 ч после заражения, отражая высокую скорость риновирусной репликации и начало выделения вируса. Главный синдром, назофарингит (собственно простуда), возникает через 1—2 дня. Он складывается из симптомов воспалительного поражения слизистых оболочек носа, рото- и носоглотки, реже — гортани. Температура — нормальная или субфебрильная, общее состояние нарушено мало. Пик приходится на 2—3-й день, средняя продолжительность заболевания составляет 7 дней. Впрочем, все показатели сильно варьируют в зависимости от серотипа, инфицирующей дозировки и особенностей организма. Симптомы могут купироваться уже через день, не получая развития, но иногда для полного выздоровления требуется более месяца.
Тяжесть и длительность заболевания коррелируют с количеством выделяемого вируса. Ограничение инфекции респираторным трактом, прежде всего его самыми верхними отделами, принципиально отличает риновирусы от других пикорнавирусов человека — энтеровирусов и вируса гепатита А, для которых характерны энтеральная передача и генерализация инфекции (вирусемия). Это можно объяснить тем, что риновирусы быстро инактивируются в кислой среде, чувствительны к желчи (отсюда неспособность к заражению энтеральным путем) и склонны к усиленной репликации при пониженной температуре, которую они находят в носовой полости.
В органных и клеточных культурах риновирусы разрушают реснитчатые эпителиоциты трахеи и носовой полости, но in vivo повреждения минимальны. Более того, даже в зонах максимального размножения (носоглотка, носовые ходы) риновирусы обнаруживаются в единичных клетках, разбросанных по слизистой оболочке (рис. 1). Это заставляет усомниться в лидерстве прямого цитопатического эффекта, побуждая думать о реактивном характере патологии. Полагают, что риновирусная инфекция служит стимулом для гиперсекреции флогогенных медиаторов и парасимпатических нейрогенных рефлексов, которые в сумме определяют основной симптомокомплекс — вазодилатацию, транссудацию сывороточных белков, гиперсекрецию слизи, приток воспалительных клеток (прежде всего нейтрофилов), чихательный и кашлевой рефлексы. Назальный секрет содержит большое количество вазотропных агентов (брадикинин, гистамин, простагландины), которые при инстилляции в нос возбуждают симптомы простуды (ринорея, заложенность носа, боль в горле). Патогенетически значимым может быть и повышение мукозальной концентрации флогогенных цитокинов (интерлейкины-1, -6, -8).

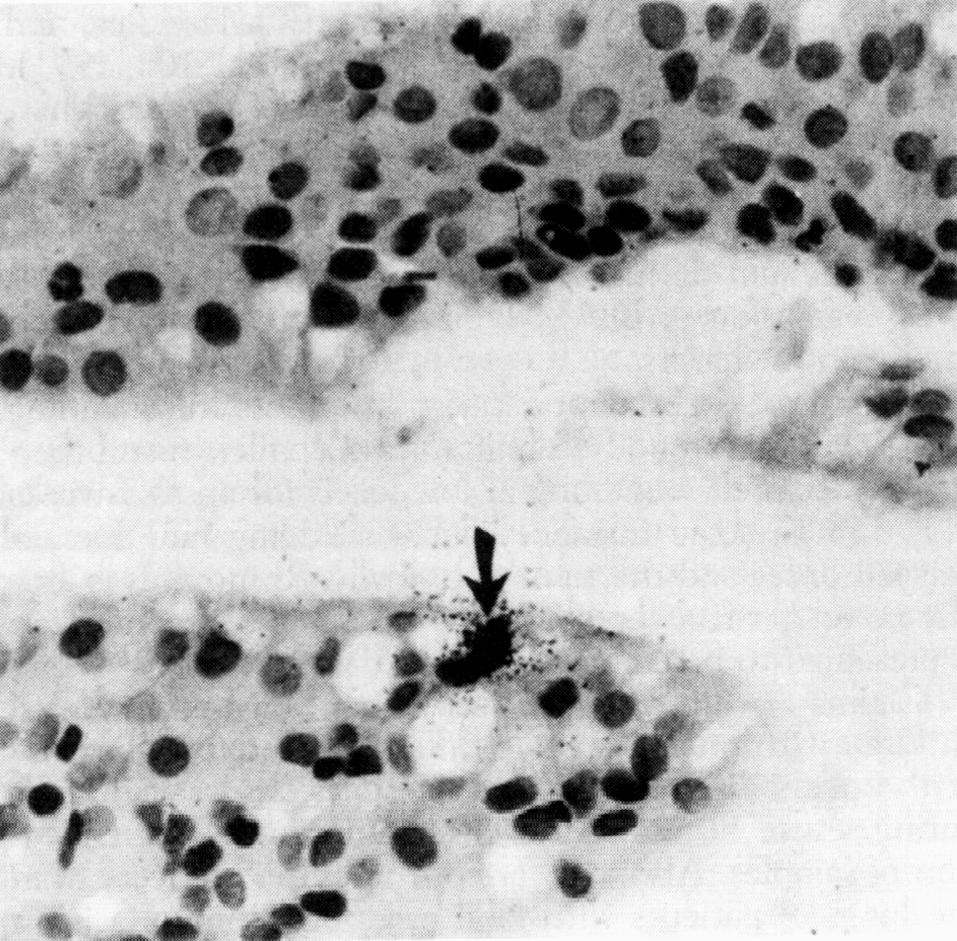
В опытах на добровольцах с экспериментальной инфекцией признаки риновирусной репликации обнаружены в бронхиальных биоптатах. Впрочем, если это и имеет значение, то скорее всего как фактор, обостряющий течение хронического бронхита и бронхиальной астмы. Не исключено, что наряду с другими респираторными вирусами риновирусы не только обостряют течение астмы, но и содействуют ее изначальному развитию, нарушая мукозальный гомеостаз бронхов.
Данные о том, что риновирусы инфицируют легкие, неубедительны. Из редких осложнений возможны синусит и средний отит. Они обычно имеют бактериальную природу, возникая из-за нарушений дренажа (отек слизистой носа!) параназальных синусов и среднего уха. Преходящие признаки синусита и среднего отита возможны и при неосложненном течении риновирусной инфекции.
Заболевание сопровождается образованием типоспецифических вируснейтрализующих антител, но их удается обнаружить не ранее 7 дней после первых симптомов. Это означает, что, как и при других ОРВИ, антитела не играют существенной роли в клиническом выздоровлении, хотя и содействуют освобождению от вируса. Сывороточные антитела сохраняются годами, титр местных IgA-антител падает гораздо быстрее. Прослеживается известная корреляция между наличием антител и устойчивостью к реинфекции, но это не гарантирует от повторного заражения даже тем же серотипом. Сравнительная ценность мукозальных и сывороточных антител не известна. Данные о значении интерферона противоречивы.
Известно множество препаратов, подавляющих репликацию риновирусов в клеточных культурах, однако ни один из них не нашел клинического применения. Новые надежды возлагают на ингибиторы пикорнавирусных протеаз. С точки зрения фармакокинетики главной проблемой является поддержание эффективной и одновременно безвредной для хозяина концентрации антивирусных агентов в зоне вирусной репликации.
Коронавирусы
Коронавирусы инфицируют широкий круг животных — птиц и млекопитающих. Они отличаются видоспецифичностью и вызывают различные заболевания у своих хозяев — от преходящего воспаления слизистых оболочек (респираторный, пищеварительный тракт) до тяжелой патологии внутренних органов (печень, почки, центральная нервная система). Это заставляет внимательно относиться и к коронавирусам человека. Не исключено, что их агрессивность не исчерпывается поражением мукозального тракта, хотя попытки выявить у них инвазивные потенции (в частности, для центральной нервной системы) не выглядят убедительно. Единственной безоговорочно признаваемой патологией человека, связанной с коронавирусами, являются острые заболевания верхнего респираторного тракта, где им, вслед за риновирусами, принадлежат лидирующие позиции. В структуре простудных заболеваний коронавирусы составляют около 15%; примерно такое число приходится на субклинические инфекции.
Коронавирусы человека были открыты в 1960-х гг., т.е. примерно через 10 лет после выделения риновирусов. Убедительные доказательства их причастности к острой респираторной патологии были получены в опытах на добровольцах, которых инфицировали назальными смывами, не содержащими риновирусов. При заражении аналогичными препаратами органных культур (трахея эмбрионов человека) удалось обнаружить вирусные частицы, похожие на уже известные вирусы инфекционного бронхита птиц и гепатита мышей. Обобщающий термин «коронавирусы» был предложен в 1968 г., название семейства, Coronaviridae, — в 1975 г.
Вирусологически это совершенно самостоятельная, даже уникальная группа, хотя с точки зрения медицинской экологии коронавирусы человека мало чем отличаются от риновирусов. И те и другие распространены повсеместно, инфицирование начинается в раннем детстве, антитела обнаруживаются почти у 100% взрослых здоровых людей. Подобно риновирусам, коронавирусы наиболее активно реплицируются при 33—34°С, что отражает их склонность к поражению верхних дыхательных путей, прежде всего слизистой оболочки носа. Вся клиническая информация о риновирусной инфекции может быть использована для коронавирусов, и мы не будем ее повторять. То же самое, по-видимому, справедливо и для патогенеза, хотя механизмы коронавирусной инфекции изучены слабо. Среди других причин это объясняется трудностями выделения коронавирусов человека: их рост в клеточных культурах наблюдается лишь после многочисленных пассажей.
Название «коронавирусы» отражает морфологические особенности вириона. Это сферические частицы размером 100—150 нм, окруженные булавовидными шипами, напоминающими солнечную корону (рис. 2). Главный поверхностный белок (точнее, гликопротеин) S (от англ. surface, поверхностный) выполняет функции рецептора, фактора слияния и протективного антигена. S-шипы отходят от липидной оболочки — суперкапсида, внутри которого находится спиралевидный нуклеокапсид. Стыковку между белком нуклеокапсида N (от англ. nucleoprotein) и суперкапсидом обеспечивает М-белок. Это необходимо для созревания, а затем для стабилизации вириона.

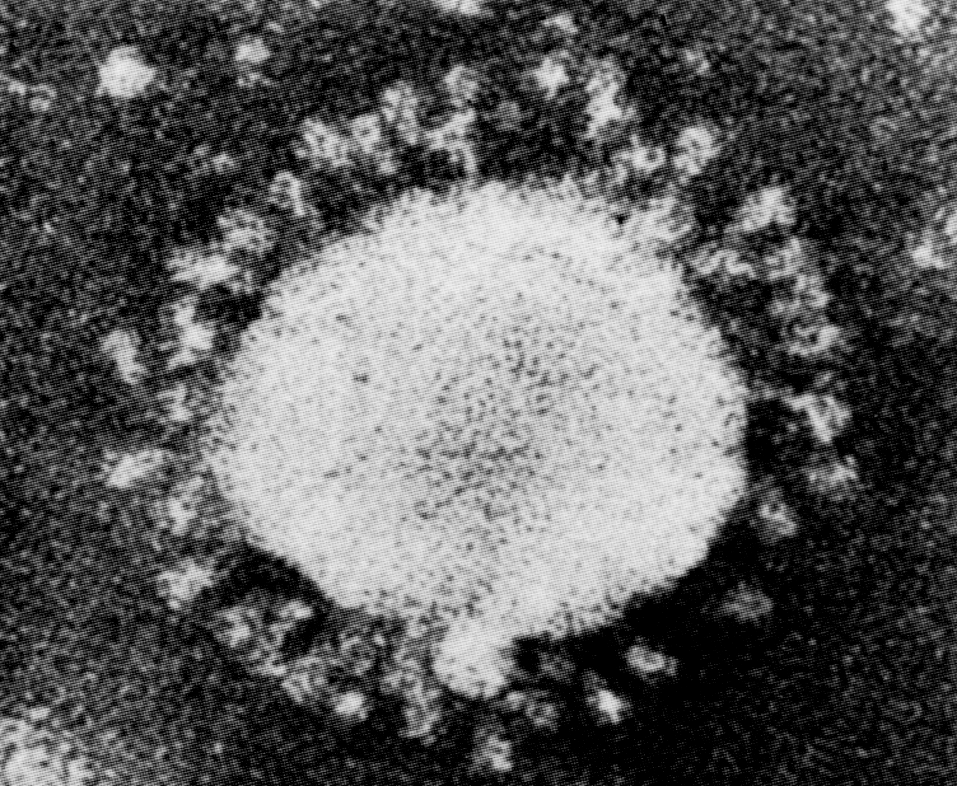
Геном представлен односпиральной несегментированной молекулой РНК, самой крупной среди вирусных РНК (20—30 кбайт, мол. масса (5—6)·106 Да). По своей стратегии она соответствует плюснитевым РНК, но репликация происходит иначе, чем у других (+)РНК-вирусов. Здесь отсутствуют два этапа, свойственные классическим (+)РНК-вирусам — синтез полипротеина и его расщепление на зрелые (структурные и неструктурные) вирусные белки. Вместо этого коронавирусы синтезируют необычную РНК-зависимую РНК-полимеразу (она образуется тотчас после проникновения вирионной РНК в цитоплазму), которая, связываясь с инициаторным (лидерным) сайтом вирусной РНК, обеспечивает образование дискретных мРНК для каждого из вирусных белков. Подобного рода механизм, сочетающийся с построением полных геномных молекул РНК, уникален для коронавирусов.
Еще одна особенность связана с образованием суперкапсида и сборкой вирионов. Эти события происходят внутри эндоплазматической сети; зрелые вирусные частицы выводятся из клетки в составе везикул по каналам эндоплазматического ретикулума и аппарата Гольджи. Плазматическая мембрана не принимает участия в образовании вирионов. Весь цикл репродукции (он начинается после рецепторзависимого слияния вирионной и клеточных мембран и проникновения вирионной РНК в клетку) происходит в цитоплазме. Ядро клетки остается интактным на протяжении всего процесса. Зараженная клетка гибнет в результате цитолиза.
Семейство Coronаviridae включает два рода — Coronavirus и Torovirus. Вирусы человека относятся к роду Coronavirus и представлены двумя видами, или скорее серологическими группами с типовыми штаммами 229Е и ОС43. Однако подобная классификация скорее всего упрощает дело. Различные изоляты могут существенно отличаться по антигенным свойствам S-протеина — главного протективного антигена коронавирусов. В ряде руководств говорится по меньшей мере о пяти серотипах, нейтрализация которых требует самостоятельных (типоспецифических) антител. Нет полной ясности и в отношении спектра коронавирусных инфекций человека. В частности, до сих пор дебатируется вопрос об энтеральных коронавирусах. Агенты, похожие на коронавирусы, находили в фильтратах фекалий новорожденных и младенцев с острым энтеритом и изредка при вспышках некротизирующего энтероколита. Сходные наблюдения есть и для взрослых, прежде всего больных СПИДом. Однако сделать окончательные выводы не удалось из-за высокого процента позитивных находок (иногда длительного носительства) у здоровых людей, особенно при низком санитарно-гигиеническом уровне. Выше говорилось и о проблеме инвазивных коронавирусных инфекций. Они хорошо документированы для животных, но их вероятность для человека еще более проблематична, чем интестинальная коронавирусная патология.
В 2003 г. случилась эпидемическая вспышка респираторной инфекции, которая из-за тяжести и количества летальных исходов получила название SARS — тяжелый острый респираторный синдром (англ. severe acute respiratory syndrome). Вспышка началась в Китае и стала быстро распространяться по другим странам. Большинство больных зарегистрировано в Китае, Вьетнаме, Сингапуре и Канаде. Всего на 29 мая 2003 г. в мире отмечено 8295 случаев SARS. От пневмонии и/или острого дистресс-синдрома погибло 750 человек. В патогенезе SARS задействованы как прямая вирусоцидность, так и иммунологические механизмы. К счастью, вспышка на этом остановилась, возможно, из-за принятых экстраординарных мер, направленных на предупреждение распространения инфекции. Оказалось, что этиологическим фактором SARS является ранее не известный коронавирус. Среди его возможных источников подозревают диких животных, в частности одну из разновидностей мангустов, которые используются в китайской кулинарии.
Специфическая терапия и вакцинация отсутствуют. Судя по опытам на добровольцах, интерферон защищает от коронавирусной инфекции. Обычным является реинфицирование штаммами той же серогруппы. Это может быть связано с быстрым увяданием мукозального иммунитета и/или заражением близкородственными, но достаточно дискретными по протективным эпитопам штаммами.
До недавних пор диагностика сводилась к анализу динамики антител, и лишь в последнее время появилась возможность обнаружения в респираторных секретах коронавирусных антигенов и РНК.
Аденовирусы
Не то я, чем кажусь…
- История открытия.
- Характеристика вириона.
- Патогенетическая неоднородность.
- Репликация: ранние и поздние белки.
- Проблемы онкогенности и персистенции.
- Аденоассоциированные вирусы.
- Аденовирусы и SV40 паповавирус.
Аденовирусы были обнаружены в начале 1950-х гг. Открытие произошло случайно, при попытке получить линии клеток из миндалин и аденоидов, удаленных у детей. После недолгих разночтений это определило название вирусов и наметило их базисную экологию, связанную с бессимптомной персистенцией в лимфоидной ткани. Но вскоре сходные инфекционные агенты были изолированы от детей и взрослых, болеющих различными респираторными заболеваниями. Интрига вокруг патогенетической функции аденовирусов обострилась после того, как были зафиксированы их туморогенные свойства — способность индуцировать злокачественные опухоли у животных (новорожденных хомячков). Онкогенный потенциал аденовирусов послужил стимулом к их тщательному изучению, оказавшемуся полезным для исследования механизмов вирусной инфекции и молекулярных процессов в эукариотических клетках.
На модели аденовирусной инфекции были впервые изучены сплайсинг, аденилирование и кэпинг матричных РНК, последовательность и регуляция экспрессии вирусных генов, их интеграция с клеточными хромосомами и пр. И хотя, к разочарованию вирусологов, оказалось, что онкогенность аденовирусов — чисто лабораторный феномен, никак не связанный с опухолями человека, аденовирусная инфекция остается важнейшим объектом молекулярной биологии и генетических манипуляций.
Термин «аденовирусы» имеет экологическую окраску, отражая склонность к персистенции в лимфоидной ткани, благодаря чему аденовирусы удается выделять из миндалин, аденоидов, аппендикса и лимфатических узлов практически здоровых людей. Но этот признак неодинаково выражен у разных аденовирусов, и не все из них могут быть отнесены к так называемым латентным разновидностям. Есть и такие, которые вызывают острые эпидемические инфекции, надолго не задерживаясь в организме хозяина.
Изучение, стартовавшее с латентных аденовирусов, быстро вышло на уровень медицинской и ветеринарной патологии, определив широкий круг частных и общих проблем, связанных с аденовирусной инфекцией. Выделено не менее 120 вирусов, которые инфицируют млекопитающих и птиц. С патогенетической точки зрения их объединяет склонность к поражению респираторного тракта, и не случайно в руководствах по инфекционным болезням аденовирусы фигурируют среди «респираторных» вирусов, т.е. возбудителей заболеваний дыхательных органов. Впрочем, к этому, в общем верному, правилу есть дополнения: аденовирусы способны вызывать поражения кишечника и конъюнктивы, а также центральной нервной системы, мочевого пузыря и половых органов.
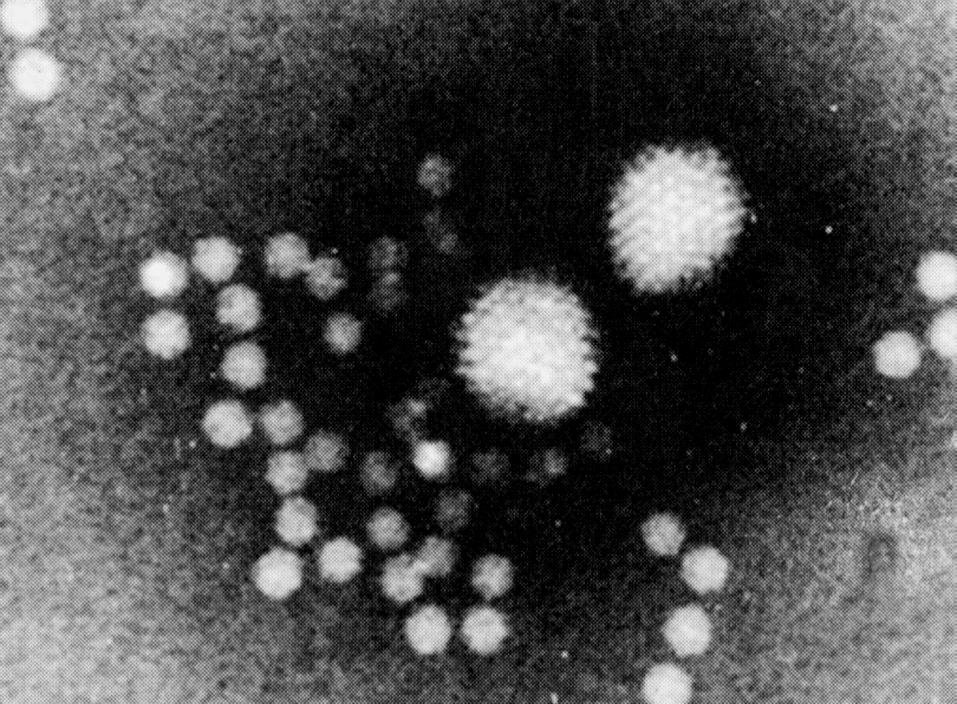
Наиболее ярким признаком аденовирусов служит морфология и структура вириона (рис. 1). Несомненно, это один из самых изящных вирусов. Вирионы лишены оболочки, имеют 80—110 нм в диаметре, капсид построен по типу кубической симметрии. В капсиде 252 капсомера: 240 из них образуют 20 равносторонних треугольных граней (икосаэдр), на углах которых расположено 12 вершинных капсомеров. Каждый из 240 капсомеров имеет шесть соседних; поэтому они называются гексонами; вершинные капсомеры соседствуют с пятью гексонными капсомерами и в связи с этим получили название пентонов. Пентоны состоят из основания и отходящей от него нити — фибриллы длиной 9—30 нм, заканчивающей утолщением — головкой величиной 4 нм. Именно фибриллы придают особую элегантность вирионам, сообщая им сходство с космическим спутником.

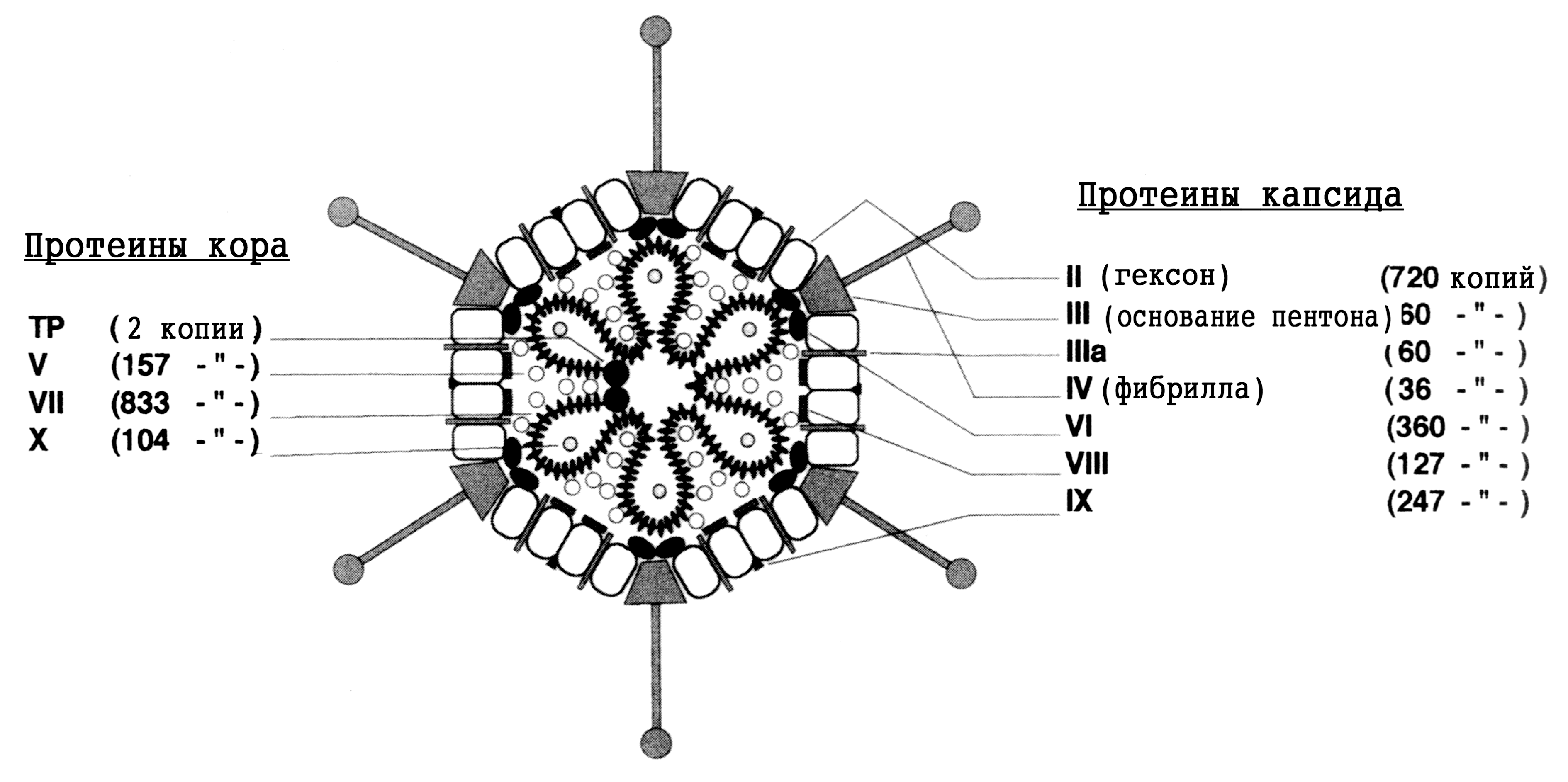
Гексоны, пентоны и фибриллы построены из разных белков (полипептидов), благодаря чему отличаются по антигенным свойствам и выполняют неодинаковые функции в инфекционном процессе. Фибриллы обеспечивают связывание с клеточными рецепторами и участвуют в дискриминации зараженных клеток, вызывая подавление синтеза клеточных макромолекул. Растворимые белки основания пентона вызывают цитопатический эффект, похожий на действие инфекционных аденовирусов, но проявляющийся гораздо быстрее (через 4—6 ч). Патогенетическое значение гексонов подтверждается тем, что антитела против их эпитопов обладают вируснейтрализующим эффектом. Возможно, они участвуют в развитии рецепторзависимого эндоцитоза, инициируемого фибриллами, — главной мишени для вируснейтрализующих антител. Полипептиды синтезируются с большим избытком: например лишь 1/10 часть молекул гексона включается в вирион.
С капсидом ассоциировано еще несколько белков, которые цементируют гексоны и структурно сопрягают капсид с сердцевиной. Последняя состоит из двухспиральной линейной ДНК, связанной с тремя полипептидами, на долю которых приходится около 20% белка вириона. Особенностью аденовирусной ДНК является наличие концевого белка — ТР (от англ. terminal protein), который ковалентно соединен с 51-концом каждой из нитей ДНК. Одной из возможных функций ТР является прикрепление ДНК к ядерному матриксу после проникновения вирусного генома в ядро. Благодаря взаимодействию концевых белков ДНК удерживается в виде кольцевой структуры, которая раскрывается при обработке протеолитическими ферментами. Так или иначе, но наличие ТР примерно в 100 раз повышает эффективность трансфекции клеток аденовирусной ДНК, изолированной из вирионов.
Размер ДНК составляет (20—25)·103 кДа, что соответствует примерно 36000 пар оснований. По логике генетического кода этого бы хватило примерно на десяток белков среднего размера. Но информационная емкость ДНК гораздо больше: аденовирусы синтезируют около 40 белков. Это достигается благодаря считыванию информации с обеих нитей ДНК и альтернативному сплайсингу, который обеспечивает получение нескольких разновидностей мРНК на основе первичного транскрипта одного гена. В зрелый вирион включается 13 белков, остальные относятся к неструктурным комопонентам, функционируя на этапе внутриклеточной репродукции вируса.
Классификация
Аденовирусы объединены в семейство Adenoviridae, которое подразделяется на два рода: Mastadenovirus (аденовирусы млекопитающих) и Aviadenovirus (аденовирусы птиц). Кроме экологического своеобразия они имеют и ряд других особенностей, которые наиболее радикально сфокусированы в антигенных свойствах вирионных белков. Аденовирусы каждого из родов имеют широкие антигенные перекресты, но лишены антигена, общего для всего семейства. Главный родоспецифический эпитоп расположен на внутренней поверхности гексона и поэтому может быть обнаружен только в растворимом гексоновом белке, например в реакции связывания комплемента (отсюда понятие «комплементсвязывающий родоспецифический антиген»).
Внутриродовая дифференцировка аденовирусов строится на разных критериях:
1. Экологические группы. Род Mastadenovirus включает вирусы (группы) человека, обезьян, крупного рогатого скота, овец, лошадей, свиней, собак, опоссума и мышей. Клинически значимых перекрестных инфекций не происходит. Более того, экологическая привязка к определенному кругу хозяев проявляется и в клеточных культурах, пермиссивность которых для аденовирусов характеризуется отчетливой видовой специфичностью.
2. Серологические типы. Их классифицируют по способности аденовирусов индуцировать нейтрализующие антитела. Серотипы аденовирусов не дают перекрестных реакций с гетерологичными вируснейтрализующими сыворотками или реагируют в минимальных титрах. Типоспецифические антигены представлены белками фибрилл и гексонов. От человека изолировано более 50 серотипов. Они объединяются в подгруппы (A, B, C, D, E, F) и играют неодинаковую роль в патологии (табл. 1). Начало делению на подгруппы положили наблюдения о характере агглютинации аденовирусами эритроцитов животных, прежде всего крыс и макак-резусов. Гемагглютинирующими свойствами обладает белок фибрилл, и такая агглютинация гасится типоспецифическими антителами.
Таблица 1. Классификация аденовирусов человека
| Подгруппа | Серотипы | Подгруппа гемагглютинации* | Онкогенность (новорожденные хомячки) | % G+C (ДНК) |
| А | 12, 18, 31 | IV | Высокая | 47–49 |
| В1 | 3, 7, 16, 21 | I | Низкая | 50–52 |
| В2 | 14, 11, 34, 35 | То же | То же | То же |
| С | 1, 2, 5, 6 | III | Нет | 57–59 |
| D | 8–10, 13, 15, 17, 19, 20, 22–30, 32, 33, 36–39, 42 | II | Нет | 57–60 |
| E | 4 | III | Нет | 57 |
| F | 40, 41 | III | Нет | 55–59 |
*Подгруппы гемагглютинации классифицируются следующим образом: I — полная агглютинация обезьяньих эритроцитов; II — полная агглютинация крысиных эритроцитов; III — частичная агглютинация крысиных эритроцитов; IV — нет агглютинации.
Исследование онкогенности вскрыло еще один аспект неоднородности аденовирусов человека: они неодинаковы по способности вызывать опухоли у грызунов (новорожденные хомячки). Удивительно, но эти, казалось бы, два совершенно разных признака (гемагглютинирующая активность и онкогенность) часто совпадают: аденовирусы, обладающие высокой и слабой туморогенностью, относятся к подгруппам с дискретным гемагглютинационным профилем (см. табл. 1).
Правомерность расчленения на подгруппы получила подтверждение в генетических исследованиях. По данным ДНК-ДНК-гибридизации, гомология оснований внутри подгрупп составляет 89—100% (исключением являются субгруппы А — 48—69% и F — 62—69%); родство между гетерологичными подгруппами не превышает 20%. Это предполагает существенные биологические различия, позволяя говорить о подгруппах как о видовых таксонах, а о серотипах — как о внутривидовых вариантах. К этому можно добавить и то, что аденовирусы различных подгрупп не рекомбинируют, хотя рекомбинации внутри группы происходят весьма эффективно. По сути, это наиболее убедительное проявление генетического родства, которое подтверждает обоснованность предложенной классификации. Согласно последним рекомендациям, именно структура ДНК, определяемая генетическими методами (гибридизация, рестрикционный анализ, секвенирование), должна быть основой систематики аденовирусов.
С практической точки зрения принадлежность к подгруппе является крупномасштабным признаком, который (пусть с изрядной долей приближения) позволяет судить об эколого-патогенетических ресурсах вирусных штаммов (табл. 2). Типовая поливалентность значительно усложняет взаимоотношения с аденовирусами, так как нейтрализация каждого серотипа требует образования гомологичных («своих») антител, не действующих на другие вирусы. Не исключено, что серотипы эволюционировали под влиянием иммунного пресса на основе случайных мутаций типоспецифических эпитопов фибрилл и гексонов. Но если это и так, то в естественных условиях процесс изменчивости идет медленно, так как антигенные свойства аденовирусов достаточно стабильны.
Таблица 2 Основная клиническая характеристика различных подгрупп аденовирусов человека
| Подгруппа | Тропизм, симптомы |
| А | Криптическая энтероинфекция |
| В1 | Респираторные заболевания |
| В2 | Почечная персистенция |
| С | Персистенция в лимфоидной ткани респираторных органов, респираторные заболевания |
| D | Кератоконъюнктивит |
| E | Конъюнктивит, респираторные заболевания |
| F | Диарея младенцев |
По крайней мере, серотипы сохраняют гомогенность на протяжении последних 50 лет, прошедших с момента открытия аденовирусов, хотя такого рода мутанты могут быть получены в лабораторных условиях гораздо быстрее. Вместе с тем принадлежность к общему серотипу не означает обязательного сходства по другим биологическим признакам. Об этом свидетельствуют генетические особенности вирусных штаммов одного и того же серотипа, выявляемые при помощи рестрикционного анализа ДНК. Это побуждает к введению категории «генотип», что, впрочем, практикуется и для ряда других вирусов, отражая стремление к максимальной конкретизации понятия о болезнетворности.
Репликация
Аденовирусы лучше всего размножаются в эпителиальных клетках естественного хозяина. Клеточные линии, полученные от животных, обычно не поддерживают инфекцию аденовирусов человека. В таких случаях дело ограничивается абортивным циклом, когда синтез ранних белков, мРНК и ДНК не завершается продукцией полного спектра капсидных компонентов. Впрочем, даже в пермиссивных системах лишь малая часть вирусных частиц обладает инфекциозностью, но огромное потомство (в каждой клетке может продуцироваться до 105—106 вирионов) обеспечивает высокую агрессивность вируса.
Репликация вирусной ДНК начинается через 8 ч после заражения и спустя 5 ч составляет около 90% от общего синтеза клеточной ДНК. Однако, несмотря на столь бурное развитие, цитопатический эффект развивается медленно и хорошо заметен лишь через 7—20 дней после заражения. Клетки округляются, отслаиваются от стекла и друг от друга, но долго не разрушаются, а вирионы (они накапливаются в ядре) не выходят в среду. В целом картина напоминает апоптоз. Среди продуктов аденовирусов есть факторы с апоптогенным эффектом (производные гена Е1А), хотя с ними конкурируют и вирусные ингибиторы апоптоза (белки, кодируемые Е1В).
Репликативный цикл аденовирусов отражает базисные механизмы продуктивной инфекции классических ДНК-вирусов. Ее принято делить на раннюю и позднюю фазы, которым соответствует экспрессия ранних и поздних генов, т.е. синтез ранних и поздних белков. Их обозначают буквами «Е» и «L» (от англ. еarly — ранний и late — поздний). Поздняя фаза берет отсчет с репликации вирусной ДНК. Она сочетается с транскрипцией поздних генов и синтезом поздних белков. Впрочем, понятия «ранний» и «поздний» не следует понимать буквально, так как для ряда процессов временные различия размыты. Многие ранние мРНК синтезируются и во время поздней фазы, а некоторые поздние белки появляются уже в начале репликативного цикла. Но в целом экспрессия генов происходит в определенной последовательности, и функции большинства ранних и поздних белков принципиально различаются между собой.
Поздние белки в основном относятся к структурным компонентам, т.е. входят в состав вириона. Ранние белки выполняют регуляторные и ферментные функции на этапах ранней и поздней транскрипции, ДНК-репликации и в посттранскрипционных событиях. Не включаясь в вирион, они играют решающую роль во взаимоотношениях вирусной ДНК с хозяином, вынуждая клетку работать на преимущественное воспроизводство вирусных белков и ДНК. В этом вирус пользуется не только своими, но и клеточными компонентами. Достаточно сказать, что, не располагая собственным инструментом транскрипции, он подключает клеточную РНК-полимеразу II. В репликации вирусной ДНК участвуют клеточные факторы, которые вместе с тремя ранними вирусспецифическими белками формируют ДНК-полимеразный комплекс. Начало ДНК-репликации знаменует наиболее драматичные взаимоотношения с клеткой. Если в ранней фазе вирусные мРНК составляют лишь 10—15% от всех мРНК, транслируемых на полирибосомах, то на позднем этапе клетка почти целиком переходит на синтез вирусспецифических белков, обеспечивая материал для мультикопирования вирионов, которое завершается гибелью зараженной клетки. Удивительно, что такого рода дискриминация совершается на фоне продолжающегося синтеза клеточных мРНК. Их выход в цитоплазму блокируют два ранних аденовирусных белка (Е1В 55 кДа и Е4 34 кДа), которые, возможно, интерферируют с транспортом мРНК из ядра.
Но мы забежали вперед — всему этому предшествует проникновение вириона в клетку, высвобождение (раздевание) вирионной ДНК и ее подготовка к транскрипции и репликации. Вирус прикрепляется к клеткам концами своих фибрилл. Природа клеточных рецепторов точно не установлена, но скорее всего это комплекс из нескольких белков, обеспечивающих мультивалентное связывание вириона с плазматической мембраной. В реакцию слияния вступают белки основания пентона, которые взаимодействуют с интегринами клеточной мембраны, приводя к эндоцитозу вирусрецепторного комплекса. В кислой среде эндосом вирион частично теряет свою капсидную оболочку и после выхода в цитоплазму при содействии микротрубочек проникает через поры ядерной мембраны в ядро, сбрасывая остаток гексонов и полностью освобождая ДНК от белков. Весь процесс раздевания вирусной ДНК занимает 1—2 ч.
Белки сердцевины замещаются клеточными гистонами с образованием хроматиноподобного комплекса, готового к экспрессии ранних вирусных генов. Среди последних есть лидер, Е1а, который транскрибируется первым и поэтому называется сверхранним. Один из трех кодируемых им белков выполняет функцию транскрипционного активатора, индуцируя транскрипцию для остальных ранних генов. Лидерству Е1а способствует его соседство с терминальным белком на конце ДНК, который служит затравкой для РНК-полимеразы. В репликации аденовирусов немало и других тонкостей, которые отражают сложные взаимоотношения не только с зараженными клетками, но и с организмом хозяина в целом. Отметим, в частности, способность аденовирусов нейтрализовать механизмы антивирусного иммунитета. Благодаря двухспиральным молекулам РНК они блокируют интерферонзависимую инактивацию фактора элонгации 2, участвующего в синтезе вирусных белков. Антиинтерфероновой активностью обладает и белок Еа1, блокирующий транскрипцию генов, индуцируемых интерфероном. Производные гена Е3 подавляют транспорт молекул главного комплекса гистосовместимости класса I (HLA-I) на поверхность зараженных клеток, тем самым защищая их от иммунного цитолиза.
Сборка вирионов происходит в ядре, куда структурные белки транспортируются из цитоплазмы после их синтеза на рибосомах и частичного объединения в капсомеры, т.е. в мультипептидные комплексы. Каждый гексоновый капсомер собирается из трех идентичных белков, пентон образуется из пяти одинаковых белков пентонового основания и трех белков фибриллы. В агрегации гексоновых субкомпонентов участвует формообразующий поздний белок (продукт гена L4), который не включается в состав вириона.
Оказавшись в ядре, капсомеры самоорганизуются в капсиды, куда включается вирусная ДНК. Конструкцию вириона завершает вирусная цистеин-протеиназа (продукт гена L3), которая доводит до финальной кондиции структурные белки. Общая продолжительность репликативного цикла составляет примерно 24 ч, но, как уже говорилось, новые вирионы не торопятся с выходом из клетки, дожидаясь ее гибели. Сборка вирионов происходит неэкономно. Структурные белки синтезируются в большом избытке, оставаясь неиспользованными. Они накапливаются в ядре в виде кристаллов вместе со скоплением зрелых вирионов (рис. 2).

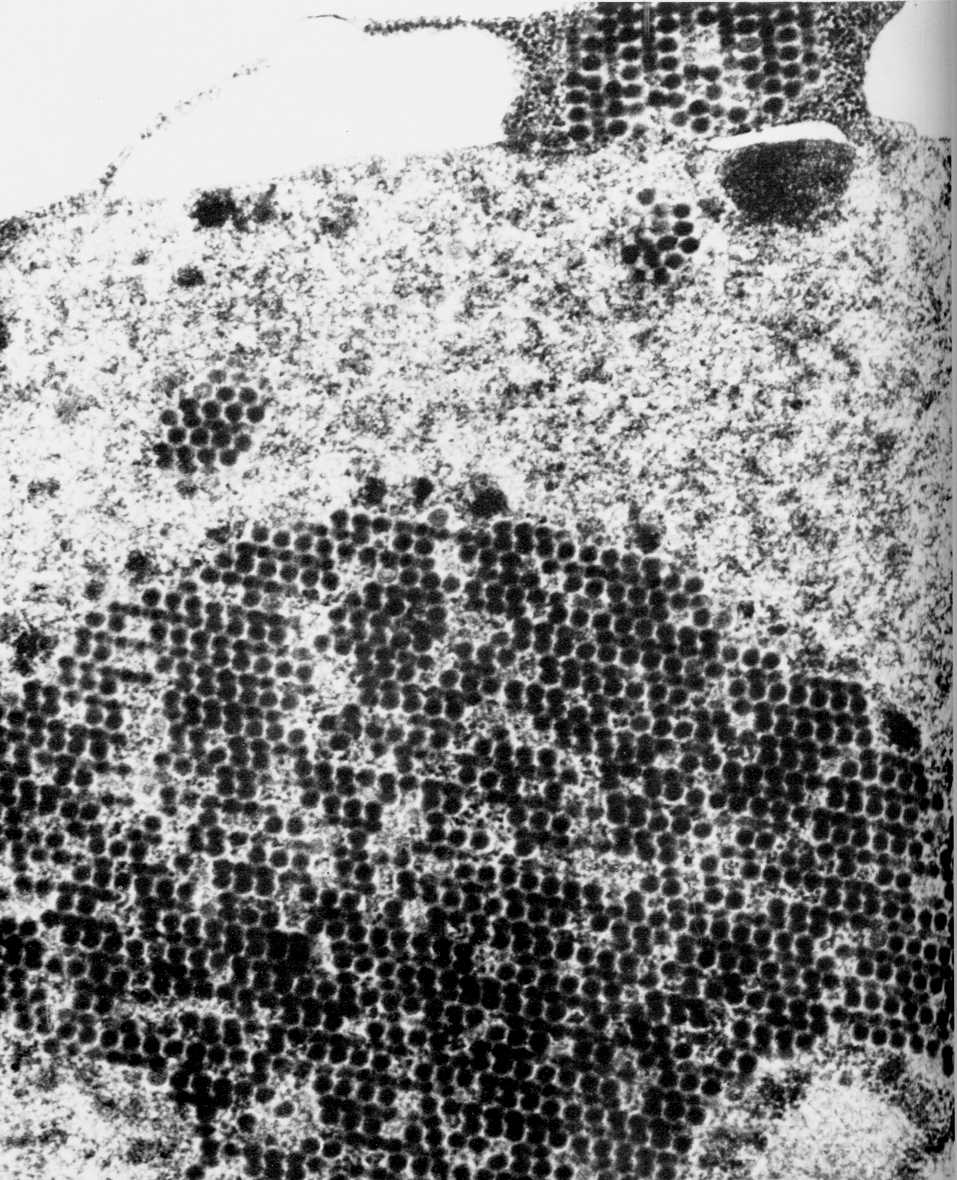
Даже такое краткое описание аденовирусной инфекции демонстрирует ряд вирусспецифических факторов и механизмов, подавление которых могло бы приостановить развитие инфекционного процесса. Однако до сего времени ни один из них не использован в качестве мишени для этиотропной терапии.
Онкогенность
Несомненно, самым интригующим фактом является способность аденовирусов оказывать туморогенное действие на клетки. Принципиально, что за исключением экспрессии нескольких ранних генов этот эффект не имеет ничего общего с продуктивной инфекцией, т.е. с воспроизводством инфекционного вируса. Более того, туморогенность наблюдается только в непермиссивных или полупермиссивных системах, т.е. по сути отражает абортивную инфекцию, при которой не происходит полной экспрессии вирусного генома, а репликативные события ограничиваются синтезом ранних белков. В таких случаях клетки не погибают, а становятся бессмертными (иммортализуются), обретая злокачественные потенции, губительные для хозяина. К сожалению (или к счастью), прагматичный финал интриги разочаровал. Аденовирусные ДНК и мРНК не удалось найти ни в одной из опухолей человека. Ничего особенного не обнаружено и иммунологами: антитела к туморассоциированным антигенам (Т-антигенам), которые являются обязательным спутником опухолевого процесса, провоцируемого аденовирусами в опытах на животных, отсутствуют у больных злокачественными новообразованиями. Это означает, что роль аденовирусов в возникновении человеческих опухолей должна быть отвергнута. Онкогенность проявляется только в модельных опытах на животных и для ее реализации требуются достаточно искусственные условия. Тем не менее, изучение аденовирусов сыграло большую роль в становлении современных концепций, а аденовирусная инфекция остается одной из главных моделей молекулярной онкологии.
Прежде всего следует отметить, что все аденовирусы человека вызывают стабильные морфофункциональные изменения в культурах непермиссивных и полупермиссивных клеток грызунов, напоминающие то, что происходит при естественном канцерогенезе. Онкологи называют это трансформацией. Трансформированные клетки размножаются быстрее нормальных, менее требовательны к питательной среде, утрачивают «чувство локтя» (т.е. выходят из-под контроля контактной ингибиции) и поэтому растут хаотично, многослойно, а главное — обретают фенотип бессмертия, т.е. способность к бесконечным инвитровым пассажам, что может быть связано с инактивацией теломеразы — фермента, укорачивающего концевые отрезки ДНК при каждом делении клеток. Центральным событием является интеграция вирусной ДНК с хозяйским геномом. Для этого требуется лишь небольшой участок (8—14%) аденовирусной ДНК, расположенный на одном из ее концов. Здесь локализованы гены Е1а и Е1b, и именно их продукты (Т-антигены) ответственны за трансформирующий эффект. Этим объясняется то, что трансформацию можно вызвать не только цельным вирусом, но и его ДНК и ее фрагментами (большинство клеток содержит не более 20% аденовирусного генома). Т-антигены содержатся в основном внутри ядра, определяя особенности трансформированных аденовирусами клеток, по которым их можно отличить от опухолевых клеток иного происхождения.
Механизм трансформации не совсем ясен. Сравнение клеточных линий показало, что вставки вирусных ДНК-фрагментов в хромосомы носят случайный характер, т.е. лишены сайтов для предпочтительной интеграции. С этой точки зрения трансформацию трудно представить как результат инсерционного мутагенеза, т.е. как дестабилизацию клеточного генома вирусными вставками. Против этого говорит отсутствие сайтовой специфичности интеграционного процесса, с чем можно было бы связать извращение функции клеточных генов (онкогенов), контролирующих пролиферацию. Скорее всего эффект опосредован через взаимодействие аденовирусных белков с регуляторами роста и деления клеток. Вряд ли случайно, что среди продуктов трансформирующей области аденовирусного генома есть факторы, влияющие на апоптоз, т.е. на генетически запрограммированную гибель клеток.
Второй аспект проблемы связан с онкогенностью in vivo. Оказалось, что некоторые аденовирусы человека вызывают опухоли у грызунов (наиболее чувствительны новорожденные хомячки). По данному признаку аденовирусы делятся на высокоонкогенные, слабоонкогенные и неонкогенные (см. табл. 1). Вряд ли можно сомневаться, что в основе онкогенности лежит эффект трансформации клеток, создающей прецедент для бесконтрольного размножения. Об этом говорит развитие опухолей после введения животным «адекватно» трансформированных клеток. Вместе с тем критерии такой «адекватности» отсутствуют, и далеко не все линии трансформированных клеток обладают туморогенностью. В частности, не выявлено очевидной корреляции между какими-либо признаками трансформированных клеток и их онкогенностью для новорожденных хомячков. Трансформацию вызывают даже неонкогенные серотипы, но опухоли надежно воспроизводятся лишь с представителями подгрупп А и В. Несомненно, что онкогенный потенциал зависит от дополнительных изменений фенотипа, и не исключено, что, несмотря на формальное сходство, трансформация, вызванная разными вирусами, несет в себе неодинаковую агрессивность.
Обращает на себя внимание тот факт, что высокая степень гомологии трансформирующих областей в пределах подгрупп А, В и С (70—90%) сочетается с низкой гомологией тех же участков ДНК для других подгрупп (1—15%). Одним из следствий этого может быть функциональная неоднозначность (в том числе онкогенность) белков, кодируемых соответствующими генами. Не случайно, по-видимому, и то, что клетки, трансформированные высокоонкогенными аденовирусами, содержат больше фрагментов вирусной ДНК и, следовательно, готовы производить более широкий спектр вирусспецифических белков. Допускают, например, что определенную роль играют факторы, ингибирующие презентацию вирусных пептидов молекулами HLA-I. Это снижает атакуемость трансформированных клеток цитотоксическими Т-лимфоцитами, способствуя их выживанию в организме. Этому соответствуют наблюдения о повышении туморогенного потенциала трансформированных клеток для иммуносупрессированных животных, к которым в известной мере можно причислить и новорожденных животных. Так или иначе, но приобретение трансформированного фенотипа следует признать необходимым, но недостаточным признаком реальной туморогенности клеток. Наиболее радикальная позиция разграничивает эти явления, подразумевая, что если для трансформации достаточно экспрессии определенной части вирусного генома, то приобретение онкогенного потенциала — результат иного (возможно, мутагенного) механизма.
Персистенция
Первые аденовирусы были выделены из миндалин и аденоидов, удаленных у детей без признаков острой инфекции. Сложилось мнение, что аденовирусы находятся в организме в латентном состоянии, инфицируя лимфоидную ткань. В принципе это оказалось верным, но обнаружилось, что по склонности к персистенции аденовирусы не однородны (см. табл. 2). Многие серотипы после острых инфекций надолго не задерживаются в организме и даже получили название эпидемических. Большинство латентных вирусов относится к подгруппам В2 и С. Они могут годами персистировать в лимфоидной ткани глоточного кольца и, по-видимому, других локализаций (например, в мезентериальных лимфатических узлах). Попадая в кишечник, аденовирусы бессимптомно реплицируются в эпителиоцитах и/или пейеровых бляшках, периодически выделяясь из фекалий.
Латенция создает вероятность эндогенных рецидивов острой инфекции и хронической гиперплазии (по сути, хронического воспаления) инфицированной лимфоидной ткани. Не исключена возможность активации вирусной инфекции в миндалинах (хронический тонзиллит), мезентериальных лимфатических узлах и аппендиксе. Аденовирусы могут активироваться на фоне иммуносупрессивной терапии и у больных СПИДом. Ряд новых серотипов (серотипы 43—47 подгруппы D) были впервые изолированы именно при СПИДе (из фекалий), позволяя думать, что, выходя из-под контроля, персистентная инфекция создает благоприятные условия для эволюции аденовирусов.
Механизм персистенции аденовирусов в лимфоидной ткани остается неясным. Скорее всего это связано с низким содержанием чувствительных (пермиссивных) клеток и очень медленной репликацией вируса в лимфоцитах, т.е. с жестко лимитированной продуктивной инфекцией. Показательно, что в опытах W. Rowe и соавт., открывших аденовирусы в 1953 г., для дегенерации культуры аденоидной ткани, связанной с размножением латентных аденовирусов, потребовалось несколько недель. Не исключена и возможность интегративной вирогении с частичной экспрессией вирусного генома. Примерно в 50% миндалин взрослых удавалось обнаружить аденовирусные антигены при отсутствии инфекционного вируса.
Вирусинфицированные клетки становятся мишенью для эффекторов иммунитета. Однако аденовирусы навязывают клеткам такие свойства, которые позволяют им избегать уничтожения или, по крайней мере, уменьшают такую возможность. Аденовирусы продуцируют факторы, которые блокируют синтез и экспрессию на поверхности клеток молекул HLA-I и тем самым подавляют презентацию вирусных антигенов, атакуемых CD8+ Т-лимфоцитами. Клетки, зараженные аденовирусами, обретают повышенную устойчивость к интерферонам и ФНО-a, мощному цитотоксическому цитокину. В обоих случаях срабатывают аденовирусные белки, интерферирующие с молекулярными механизмами, которые определяют антивирусные эффекты. Среди продуктов ранних аденовирусных генов есть факторы, задерживающие развитие апоптоза. В целом это отражает стратегию, нацеленную на повышение выживаемости зараженных клеток и создание условий для вирусной персистенции.
Патогенез и клиническая агрессивность
Большинство аденовирусных инфекций протекает бессимптомно, и в таких случаях только наличие специфических антител говорит о cостоявшемся конфликте с хозяином. Но возможны и клинические проявления, которые чаще всего связаны с острым воспалительным поражением слизистых оболочек респираторного тракта, конъюнктивы и кишечника. Обычно это самокупирующиеся процессы, не требующие врачевания, хотя изредка возникают непредсказуемые осложнения. Аденовирусы реплицируются в эпителиальном покрове слизистых оболочек и редко проникают дальше регионарных лимфатических узлов. Лимфоидная ткань активно реагирует на внедрение вируса и после выраженной воспалительной реакции (она проявляется в виде регионарной лимфоаденопатии), становится резервуаром персистентной инфекции.
Аденовирусы, болезнетворность которых доказана, проявляют ее неравнозначно (см. табл. 2). Без большой натяжки можно говорить о полиэтиологичности аденовирусных инфекций, подразумевая, что разные серотипы имеют неодинаковые клинико-эпидемиологические характеристики. Действительно, некоторые из них преимущественно выделяются при респираторных заболеваниях детей или взрослых, при поражениях конъюнктивы или острых кишечных инфекциях и пр. Многие серотипы до сих пор не удалось связать с какой-либо патологией; скорее всего они способны лишь к латентной персистенции. Из шести недавно открытых серотипов (42—47) пять явились неожиданными изолятами из фекалий больных СПИДом, а один (42) — из испражнений здорового ребенка. Их происхождение и клиническое значение не известны, но это дает повод для размышлений о мутациях и/или рекомбинациях, генерирующих обновленные варианты. Словом, есть очевидные основания для патогенетической классификации аденовирусов, которая, несмотря на определенную расплывчатость, дает представление об их клинической неоднородности.
По приблизительным подсчетам аденовирусные инфекции составляют 5—10% от всех острых вирусных заболеваний. Большинство этих инфекций падает на детский возраст; 75% возникают в возрасте до 14 лет и более половины — в возрасте до 5 лет. Аденовирусы традиционно относят к респираторным вирусам, так как их латенция и клиническая значимость с самого начала ассоциировались с респираторным трактом. Теперь мы знаем, что это неверно. Во-первых, «респираторные» аденовирусы часто выделяются из фекалий, что скорее всего связано с вторичной (как правило, бессимптомной) инфекцией энтероцитов и лимфоидной ткани кишечника. Во-вторых, есть аденовирусы (главным образом серотипы 40 и 41), наделенные избирательной энтеропатогенностью; они служат одной из причин острых кишечных заболеваний детского возраста. Наконец, не следует забывать и об аденовирусной экзотике (острый геморрагический цистит, поражения центральной нервной системы, половой сферы и пр.), которая оттеняет патогенетическую многоликость аденовирусов. Впрочем, об этом уже говорилось выше.
Аденовирусные инфекции верхних отделов респираторного тракта чаще всего возникают в раннем детстве и относятся к серотипам 1, 2, 5, 6 подгруппы С. Заболевания обычно наблюдаются в зимние месяцы и по клинике не отличаются от банальной простуды, т.е. от большинства других острых респираторных вирусных инфекций. Превалируют симптомы острого поражения верхних дыхательных путей (фарингит, насморк, кашель) и умеренные признаки общей интоксикации (лихорадка, озноб, головная боль, миалгия). Инфекция нередко переходит в латентную форму, поддерживая иммунитет против повторного заражения. К юношескому возрасту большинство имеет нейтрализующие антитела против данных серотипов, хотя уже к пятилетнему возрасту у многих имеются иммунные маркеры. Если понятие о нормальной вирусной флоре справедливо, то аденовирусы подгруппы С близко подходят к этой категории.
Остальные потенциально болезнетворные аденовирусы ведут себя более агрессивно. Они не склонны к персистенции и иногда называются даже эпидемическими. Это предполагает вероятность вспышек, но скорее подчеркивает облигатность экзогенного заражения.
Серотипы 3 и 7 (подгруппа В) служат причиной фарингоконъюнктивальной лихорадки — наиболее очерченной формы аденовирусной патологии, для которой характерна триада симптомов: конъюнктивит, фарингит, лихорадка. У взрослых (они обычно заражаются при контактах с больными детьми) процесс может ограничиться поражением конъюнктивы. Болезнь проходит без тяжелых последствий, но доставляет немало тревог, тем более что у детей иногда возникают фолликулярные и пленчатые формы конъюнктивита, похожие на хламидийную инфекцию. Нередко заражение происходит при купании (через воду). С этим связаны вспышки так называемого бассейнового конъюнктивита, который чаще всего поражает детей в летние месяцы. Те же аденовирусы ответственны примерно за 10% детских пневмоний с немалой (8—10%) вероятностью летального исхода у детей первого года жизни.
Более серьезной офтальмопатологией, которая может осложниться помутнением роговицы и нарушением зрения, является кератоконъюнктивит (серотипы 8, реже 19, 37 — подгруппа D). Он наблюдается у лиц, занятых на производствах с высоким пылевым загрязнением, создающим опасность раздражения роговицы. Заражение происходит при оказании медицинской помощи через инфицированные инструменты, глазные примочки и пр.
Аденовирусная инфекция была обнаружена во время эпидемической вспышки острой респираторной патологии среди солдат-новобранцев (США). Ретроспективный анализ (по данным иммуносерологии) и последующие наблюдения утвердили этиологическую значимость аденовирусов при острых респираторных заболеваниях среди формирующихся коллективов на фоне стрессорных воздействий и уплотненных контактов. Логика очевидна: стресс и изнуряющие тренировки снижают устойчивость к инфекции, а скученность повышает вероятность взаимозаражения. Обычно находят серотипы 4 (подгруппа Е) и 7, реже 14 и 21 (подгруппа В). Проблема оказалась настолько серьезной, что заставила задуматься над созданием специфической вакцины. Вирусы, инактивированные формалином, оказались малоэффективными, но хороший результат получен с живыми вирусами типов 4 и 7. Пожалуй, это единственный пример, когда в качестве вакцины используются не аттенуированные штаммы, а те же дикие вирусы, которые вызывают заболевание. Хитрость в том, что вакцину принимают per os в защитных (желатиновых) капсулах. Это исключает заражение фарингеального эпителия, обеспечивая высвобождение вирусов в тонком кишечнике, где возникает субклиническая инфекция с развитием протективного иммунитета против повторного респираторного заражения.
Многие аденовирусы реплицируются в эпителии кишечника и выделяются с испражнениями. Они не имеют отношения к острым кишечным заболеваниям и могут рассматриваться как элемент нормальной микрофлоры кишечника. Тем не менее, настойчивые попытки доказать причастность аденовирусов к патологии пищеварительного тракта в конце концов увенчались успехом. Известны два серотипа подгруппы F (40 и 41), которые с полным основанием могут называться кишечными аденовирусами. С ними связано 5—15% острых диарей младшего детского возраста — показатель, уступающий только ротавирусам. Экологическую специфику (т.е. тканевый тропизм) данных серотипов подчеркивает тот факт, что в отличие от других аденовирусов они с трудом поддаются культивированию, в частности не инфицируют стандартные линии клеток, HeLa и Нер2. Их в изобилии обнаруживают при помощи электронной микроскопии или иммунодиагностических тестов в фекалиях инфицированных детей.
Следует прокомментировать еще одно наблюдение, которое широко обсуждалось в связи с болезнетворностью аденовирусов. В препаратах аденовирусов иногда обнаруживаются мелкие (20 нм) вирионы, не имеющие ничего общего с аденовирусами (рис. 3). Оказалось, что это дефектные парвовирусы, которые не способны реплицироваться без участия аденовирусов. Последние выступают в роли помощников, одалживая сателлитным вирусам (их называют аденоассоциированными вирусами, AAV) некоторые из своих ранних белков. Без такой поддержки AAV-вирусы не дают потомства. Доказательств болезнетворности AAV-вирусов не получено. Более того, подавляя репликацию аденовирусов, они ведут к снижению их инфекциозности.

Любопытно, что и сами аденовирусы могут выступать в роли сателлитов. Это наблюдается при культивировании некоторых человеческих серотипов в клетках почек обезьян. Их репродукция происходит только в присутствии знаменитого обезьяньего паповавируса SV40. Это случайное открытие повлекло за собой множество работ по изучению механизмов хелперного эффекта SV40 и явилось основанием запретить получение вакцинных штаммов аденовирусов в культурах обезьяньих клеток. Дело в том, что SV40 вызывает злокачественную трансформацию клеток, и присутствие его следов в вакцинных препаратах рассматривается как угроза туморогенного действия in vivo. Впрочем, ретроспективный анализ широкого применения таких вакцин не выявил каких-либо осложнений.
Иммунология и лабораторная диагностика
Аденовирусы вызывают полный спектр иммунных реакций, но основное внимание сконцентрировано на гуморальном ответе. Протективный эффект сывороточных антител впервые обнаружен в опытах на добровольцах в 1956 г., т.е. вскоре после открытия аденовирусов. Дальнейшие исследования подтвердили это наблюдение, но сделали его более прицельным: вируснейтрализующей активностью (и, соответственно, протективностью) обладают антитела к типоспецифическим антигенам; антитела против группового (общего для всех аденовирусов человека) антигена не эффективны. Принципиально, что сывороточные антитела гарантируют высокий уровень защиты при отсутствии местных антител. Этот явный диссонанс с другими респираторными вирусами подчеркивает своеобразие аденовирусной инфекции, прежде всего патогенетическую значимость инвазивного компонента. Речь идет о вирусемии, которая так часто сопутствует аденовирусной инфекции, что иногда рассматривается как ее фаза. Характерным является повреждение сосудистого эндотелия, которое содействует выходу плазменных факторов (в том числе антител) в зону поражения. Циркулирующие антитела способны оборвать развитие каждого из этих процессов.
Типоспецифический иммунитет отличается продолжительностью, спектр его расширяется с возрастом, но не охватывает всех серотипов, оставляя лазейку для новой аденовирусной патологии. Циркулирующие антитела более надежно защищают от клинически значимой инфекции, чем от повторного заражения. Это определяет вероятность бессимптомного инфицирования, поддерживая напряженность специфического иммунитета на протяжении всей жизни. Новорожденные получают антитела от матери, которые до года обеспечивают защиту от аденовирусов (опять-таки не от всех!) примерно половины младенцев. В возрасте 6—15 лет нейтрализующие антитела к серотипам 1, 2 и 5 (подгруппа С) имеют 40—60% детей, но антитела против более агрессивных аденовирусов (серотипы 3, 4 и 7 — подгруппы В и Е) выявляются реже.
Решающая роль циркулирующих антител в защите от аденовирусов не исключает влияния Т-эффекторов. Об этом говорит частое выделение аденовирусов (в том числе ранее неизвестных серотипов — см. выше) от больных ВИЧ-инфекцией. В ряде случаев это сочеталось с летальным исходом. Основной мишенью для Т-лимфоцитов являются консервативные (общие для всех аденовирусов) эпитопы структурных белков. Этим можно объяснить наблюдения о гиперреактивности Т-клеток к аденовирусным антигенам при отсутствии типоспецифических антител. Комментируя значение клеточного иммунитета, следует помнить о механизмах, позволяющих аденовирусам блокировать или хотя бы ослаблять эффекторную отдачу Т-зависимых реакций.То же самое справедливо и для интерферона, хотя некоторые клиницисты отстаивают его эффективность при аденовирусных кератоконъюнктивитах.
Мы уже говорили о вакцинопрофилактике аденовирусной инфекции во вновь организуемых воинских коллективах. Попытки воспроизвести нечто подобное в общей (гражданской) популяции не известны. Считают, что это слишком дорогостоящее и трудоемкое мероприятие, не совместимое с малым риском клинически значимых, тем более тяжелых аденовирусных инфекций.
Известен ряд препаратов, которые угнетают рост аденовирусов in vitro, но эффективной химиотерапии аденовирусных заболеваний не существует.
Классический метод диагностики аденовирусных инфекций — выделение вируса в культуре индикаторных клеток (таких как Hela, Hep2, KB, A549). Однако это неприемлемая для практики, трудоемкая и длительная процедура (цитопатический эффект заметен через 1—4 нед). Поэтому, как и при других вирусных инфекциях, чаще пользуются выявлением аденовирусных антигенов и фрагментов ДНК. В последнем случае применяют полимеразную цепную реакцию с праймерами для участка ДНК, кодирующего группоспецифический фрагмент гексона. Это дает возможность быстро, с высокой чувствительностью и унифицированно выявлять все серотипы аденовирусов человека. Спецификация праймеров позволяет дифференцировать разные серотипы. Такие тесты разработаны, в частности, для энтеропатогенных аденовирусов серотипов 40 и 41.
Достоверным диагностическим критерием является нарастание титров антител против аденовирусных антигенов. Это фиксируют в реакции связывания комплемента и/или в реакциях нейтрализации биологической активности вирусов (торможение гемагглютинации и подавление цитотоксичности в культуре клеток). Применяется редко.
Папилломавирусы
Уснула щука, да зубы не спят.
- Классификация и вирусология.
- Генотипы и патология.
- Эпидемиология папилломатоза.
- Репликативная стратегия и патогенез.
- Механизмы онкогенной трансформации эпителиоцитов.
Папилломавирусы являются родовым таксоном (Papillomavirus) семейства Papovaviridae. Папова — акроним трех групп вирусов: папилломавирусы, полиомавирусы и вакуолизирующие вирусы. В медицинской вирусологии главную роль играют папилломавирусы. Они настолько своеобразны, что масштаб родового ранга для них кажется недостаточным. Поэтому в последних руководствах о папилломавирусах говорят как о подсемействе Papovaviridae и даже самостоятельном семействе Papillomaviridae.
Папилломавирусы представлены многочисленной группой родственных вирусов, включающей патогены человека и большинства позвоночных. Для них характерна видоспецифичность, т.е. ограниченный круг поражаемых хозяев. Человек инфицируется собственными папилломавирусами, и все ассоциированные с ними болезни относятся к антропонозам.
Папилломавирусы человека (HPV, от англ. human papillomaviruses) относятся к онкогенным вирусам, т.е. вызывают образование опухолей — от безобидных до смертельно опасных. Онкогенный эффект связан с их способностью нарушать дифференцировку и индуцировать пролиферацию эпителиоцитов кожи и слизистых оболочек. Это проявляется в виде папиллом (бородавок) разного типа и разной локализации, а также эпителиальных дисплазий, которые могут трансформироваться в инвазивные (раковые) опухоли.
Кожные бородавки чаще возникают у детей и обычно персистируют несколько лет, причиняя главным образом косметические неудобства. Гораздо серьезнее генитальные (или аногенитальные) бородавки. Они называются остроконечными кондиломами (condyloma acumanita), образуя разрастания, которые в типичном виде похожи на цветную капусту. Генитальные бородавки чаще развиваются на наружных половых органах, но могут поражать влагалище и шейку матки. Это одна из самых распространенных инфекций, передаваемых половым путем.
Сходные поражения встречаются и в других отделах слизистых оболочек — ротовой полости, конъюнктиве, верхних дыхательных путях. HPV-инфекция гортани изредка служит причиной рецидивирующего респираторного папилломатоза (ларингеальные бородавки). Он обычно возникает в раннем детстве, являясь результатом перинатального заражения (генитальные бородавки). При распространении на глубокие отделы респираторного тракта к охриплости присоединяются симптомы асфиксии, требующие трахеотомии. Операцию часто приходится делать многократно.
Дисплазии (син. интраэпителиальные неоплазии) наиболее типичны для эпителия шейки матки. В большинстве случаев они носят доброкачественный характер, но возможна и малигнизация вплоть до инвазивного рака. По летальности цервикальная карцинома занимает второе место среди раковых заболеваний женщин (после рака грудной железы), а в развивающихся странах занимает первую позицию (около 25% женских злокачественных опухолей).
Вирусология
Папилломавирусы представляют собой небольшие (55—60 нм в диаметре) безоболочечные вирусы. Капсид кубической симметрии содержит два белка — L1 и L2 (от англ. late, поздний — см. ниже). L1 называется главным капсидным белком. Он составляет более 80% капсидного материала, образуя блоки (капсомеры), из которых построен капсид. Анти-L1-антитела обладают вируснейтрализующей активностью, подчеркивая значимость L1 в инициации инфекции. L2 является минорным белком. Он не задействован в образовании капсида, но участвует в его стабилизации и стыковке с геномом.
Геном представлен циркулярной двухспиральной ДНК, ассоциированной с клеточными гистонами. Он состоит примерно из 8 000 пар оснований и кодирует около 10 белков. Транскрипции подвергается только одна из нитей ДНК. Функционально геном включает три участка:
- Некодирующий (нетранслируемый) фрагмент — длинный контролирующий участок (LCR, long control region; 400—1000 пар оснований). Он управляет экспрессией вирусных генов, воспринимая (связывая) вирусные и клеточные регуляторы транскрипции. Здесь же находится участок, с которого начинается репликация ДНК. LCR — наиболее вариабельный фрагмент вирусного генома. Не исключено, что именно его особенности определяют тканевую специфичность папилломавирусов.
- Гены, кодирующие ранние белки: Е1, Е2, Е4, Е5, Е6 и Е7 (от англ. early — ранний; е3 не экспрессируется — псевдоген). Это неструктурные белки (не входят в состав вириона), задействованные в вирусной репликации и онкогенезе.
- Гены, кодирующие структурные (вирионные) белки, L1 и L2.
Классификация, репликация и патогенез
HPV делится на типы, точнее генотипы, которые дифференцируют по особенностям нуклеотидной последовательности небольшого участка l1-гена: гомология не должна превышать 90%. Внутри типов дифференцируют субтипы: их сходство (по тому же фрагменту ДНК) составляет 90—95%. Известно более 200 типов, около сотни изучено достаточно подробно.
Типирование позволяет судить о патогенетическом потенциале папилломавирусов, являясь ориентиром при оценке тканевого тропизма (кожные и мукозальные HPV-типы) и онкогенной агрессивности. Типы 1—4 обычно вызывают кожные бородавки. При аногенитальных поражениях доминируют типы 6, 11, 16, 18, 31, 33 и 35. HPV-1 чаще ассоциирован с плантарными (подошвенными) бородавками, тогда как для HPV-2 более характерны обычные (вульгарные) бородавки кистей рук.
Степень онкогенной агрессивности определяет вероятность злокачественной трансформации инфицированных клеток. HPV относятся к немногим вирусам человека, участие которых в канцерогенезе не только постулировано, но и доказано. Злокачественные опухоли, ассоциированные с HPV, могут иметь разную локализацию, но безусловное лидерство принадлежит цервикальной HPV-инфекции, за которой признается ведущая роль в развитии рака шейки матки. Из 30 HPV-типов, распространяющихся половым путем и первично инфицирующих половые органы, всего четыре типа (16, 18, 31 и 45) отвечают за 80% случаев цервикального рака. По степени корреляции со злокачественными неоплазиями цервикального канала предложено дифференцировать две группы папилломавирусов — низкого и высокого риска. Фактически это единственная реальная основа для прогнозирования злокачественной эволюции HPV-инфекции.
Наконец, HPV-типирование служит инструментом эпидемиологического анализа, позволяя следить за распространением инфекции и дифференцировать эндогенные рецидивы от реинфекции. Это позволило связать респираторный папилломатоз с перинатальным HPV-заражением и орогенитальными половыми контактами, обосновать возможность неполового пути передачи генитальных HPV-вирусов, проследить за этиологической преемственностью неоплазий и инвазивной карциномы шейки матки.
При кожных формах заражение происходит путем прямых и опосредованных контактов, т.е. через объекты внешней среды. Об этом говорят вспышки папилломавирусных инфекций (бородавок) при пользовании общими гимнастическими снарядами и посещении бассейнов. Наиболее чувствительны дети раннего возраста.
Генитальные бородавки передаются половым путем и принадлежат к числу самых распространенных сексуально-трансмиссивных заболеваний. Впрочем, возможны и неполовые пути заражения. На это указывает отсутствие корреляции между HPV-типами половых партнеров и обнаружение HPV-ДНК у девственниц. Генитальный тракт служит источником заражения ротовой полости и верхних дыхательных путей. Это происходит при перинатальном инфицировании или в результате орогенитальных половых контактов.
Папилломавирусы инфицируют многослойный плоский эпителий, что определяет спектр поражаемых мишеней — эпидермис кожи и слизистые оболочки, покрытые эпителием такого типа. Заражению подвергаются клетки базального (зародышевого) слоя. Это недифференцированные, активно пролиферирующие клетки, которые обеспечивают непрерывное обновление эпителиального пласта. Дозревая по мере продвижения на поверхность, они утрачивают способность к делению, и в конце концов сбрасываются в окружающую среду. В эпидермисе это сочетается с ороговением, т.е. превращением клеток в кератиновые чешуйки, лишенные ядра и цитоплазматических органелл.
Особенность (возможно, уникальность) папилломавирусов состоит в том, что созревание вирионов зависит от дифференцировочного процесса клеток (см. рисунок). В зародышевом слое вирус лишь закрепляет свой генетический материал, который реплицируется синхронно с хромосомами и передается в виде эписомных копий дочерним клеткам. Синтез вирусных белков начинается в шиповидных клетках, а сборка вирионов (она происходит в клеточном ядре) достигает пика в зернистом слое эпителия.

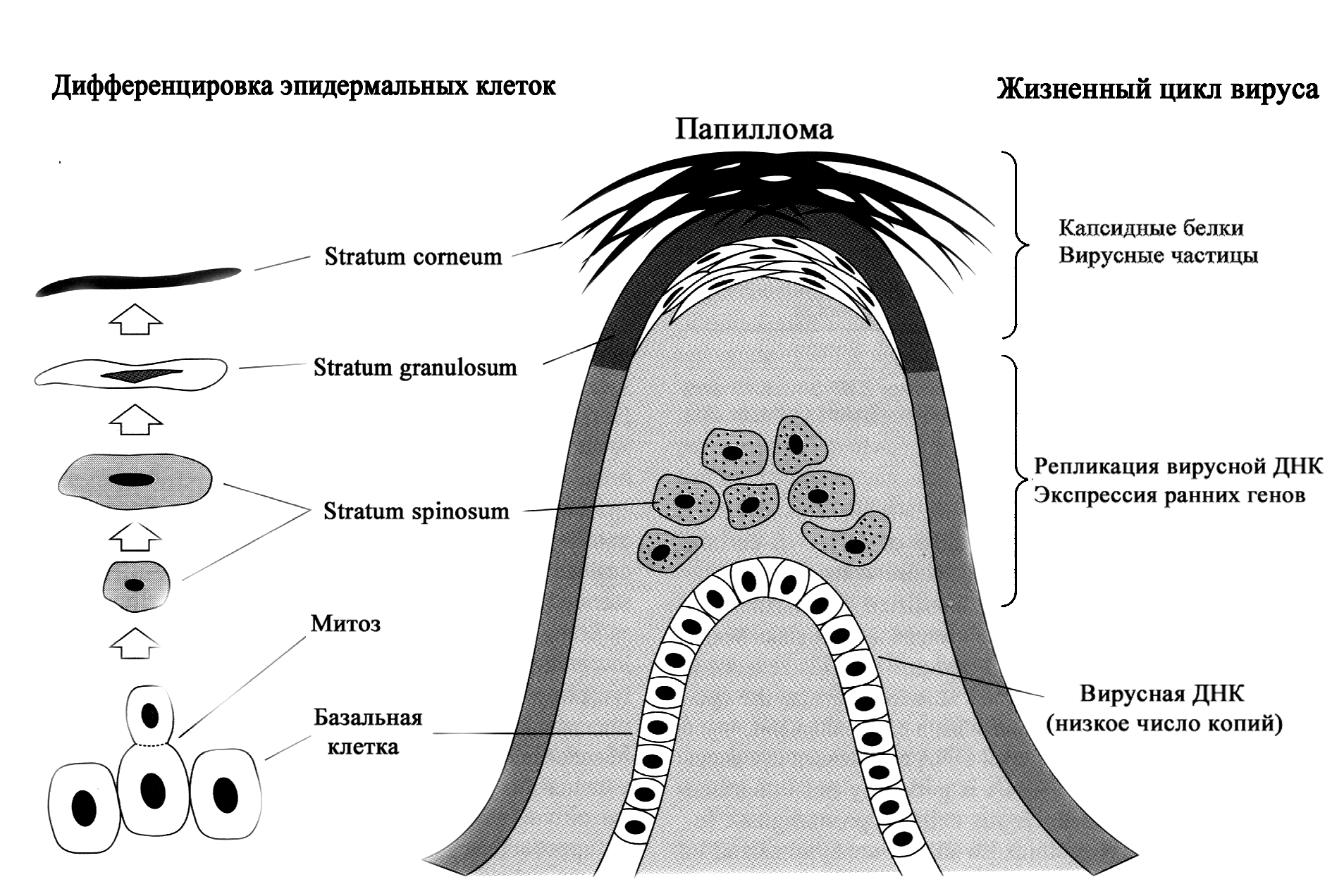
Принципиально, что речь идет о зависимости продуктивной репликации именно от дифференцировочного процесса, а не от степени зрелости клеток как его промежуточного или финального этапа. Этим, в частности, объясняются трудности культивирования папилломавирусов. Нужны не просто дифференцированные эпителиоциты, а клетки, находящиеся в процессе дифференцировки, причем заражение должно произойти на фоне высокой пролиферативной активности клеток. Только в таких условиях удается поддерживать продуктивную инфекцию папилломавирусов в клеточных культурах in vitro.
Вирус достигает базального слоя эпителия через микротравмы эпидермиса или слизистых оболочек. К числу вероятных клеточных рецепторов относятся гепарансульфат и a6-интегрин, хотя разные HPV-типы в этом отношении не одинаковы. Оказавшись в клетке, вирусная ДНК транслоцируется в ядро, где происходят основные события. Как уже говорилось, продуктивная репликация возможна лишь в клетках, вступивших на путь необратимой дифференцировки. В противном случае инфекция принимает абортивный характер. Вирусный геном элиминируется или начинает персистировать, используя механизм неинтегративной и/или интегративной вирогении; последнее характерно для малигнизированных клеток (см. ниже).
Зависимость от факторов дифференцирующихся, но утративших способность к пролиферации эпителиоцитов ставит перед вирусом непростую задачу. Для репродукции ему необходимы транскрипция и репликация ДНК, которые целиком зависят от пролиферативного потенциала клетки-хозяина. Но теряя митотическую активность, эпителиоциты прекращают синтез ферментов, поддерживающих пролиферацию. Вирус решает эту проблему, навязывая клетке пролиферативную активность, т.е. возвращает ее способность к образованию необходимых для себя транскрипционных и репликативных факторов. В этом суть репликативной стратегии и причина онкогенности папилломавирусов.
Сочетание клеточной пролиферации с дифференцировочным процессом характерно для доброкачественных опухолей (бородавки). Малигнизация останавливает процесс созревания эпителиоцитов, наделяя их злокачественной (инвазивной) агрессивностью. Если при доброкачественном росте пролиферирующие клетки не выходят за пределы эпителия (интраэпителиальный рост), то злокачественная дисплазия сочетается с пенетрацией базальной мембраны, знаменуя начало метастазирования.
Ранние HPV-белки обладают полифункциональностью. Она основана на их способности к взаимодействию с различными клеточными белками и регуляторными сайтами HPV-генома (LCR). Вопреки нумерации, репликация начинается с образования белков Е6 и Е7. Это происходит под влиянием клеточных факторов, которые связываются с LCR, запуская транскрипцию вирусных генов. Белки Е6 и Е7 реанимируют клеточный цикл, побуждая к синтезу факторов, необходимых для транскрипции и репликации ДНК. Главными мишенями служат два ключевых противоопухолевых белка, р53 и р105-RB. Они контролируют деление клеток, останавливая митотический цикл в G1-фазе. Е6 взаимодействует с р53, делая его чувствительным к убиквитинзависимой деградации. Кроме того, Е6 связывается еще с несколькими белками, функции которых не известны . Е7 действует через рRB, точнее через семейство RB-белков. Комплексирование с рRB ведет к высвобождению транскрипционного фактора E2F-1. Он обеспечивает экспрессию генов, продукты которых необходимы для S-фазы клеточного цикла. Кроме того, Е7 взаимодействует с циклинами, циклинзависимыми киназами и другими белками, регулирующими клеточный цикл. Еще одним фактором, способствующим пролиферации клеток, является белок Е5. Он синтезируется вслед за Е6 и Е7, усиливая экпрессию рецепторов для ростстимулирующих цитокинов. Это повышает чувствительность к митогенным и дифференцировочным стимулам, которые эпителиоциты получают из окружающей среды.
Обобщая, можно сказать, что благодаря взаимодействию е6-, е7- и е5-генов клетки обретают пермиссивность, т.е. способность поддерживать репродукцию HPV. В репликации HPV-ДНК участвуют белки Е1 и Е2. Е2 обеспечивает связывание инициаторного LCR-сайта с Е1, а тот в свою очередь подключает комплекс клеточных белков, необходимых для репликации ДНК. Е2 выступает и в роли регулятора транскрипции вирусных генов, определяя место сборки и действия РНК-полимеразы.
Важной функцией белка Е2 является подавление транскрипции е6- и е7-генов, т.е. синтеза белков Е6 и Е7. Это восстанавливает антипролиферативную активность р53 и рRB, содействуя дифференцировочному процессу эпителиоцитов и синтезу поздних HPV-белков, L1 и L2. Фактор, инициирующий транскрипцию l1- и l2-генов, не известен, но он, безусловно, связан с дифференцировкой эпителиальных клеток, так как образование L1 и L2 начинается по достижении ими определенной степени зрелости (в эпидермисе — клетки верхнего шиповидного и зернистого слоев (см. рисунок).
Белки L1 и L2 транспортируются в ядро, где происходит сборка вирусных частиц. Вирионы высвобождаются пассивно (без цитолиза) при дезинтеграции поверхностных клеток, возможно, при участии белка Е4. Слущиваемые клетки (в том числе роговые чешуйки эпидермиса) контагиозны.
Малигнизация. Взаимоотношения с HPV могут выходить за рамки доброкачественного конфликта, завершаясь злокачественной трансформацией эпителиальных клеток. Классический пример — рак шейки матки (цервикальная карцинома), хотя известны и другие раковые опухоли, чаще урогенитальной сферы, ассоциированные с папилломавирусами. Из 30 HPV-типов, которые распространяются половым путем и первично инфицируют урогенитальные органы, при цервикальном раке чаще всего встречаются типы 16, 18, 31 и 45. Около половины случаев ассоциированы с HPV-16, 25—30% — с HPV-18, 31 и 45. В целом группу высокого риска составляют более десятка HPV-типов. С ними ассоциировано почти 100% случаев плоскоклеточного рака шейки матки. Для цервикальных аденокарцином корреляция не столь значительна и существенно зависит от возраста. У женщин моложе 40 лет HPV присутствуют в 89% аденокарцином, у женщин 60 лет и старше — в 43%.
Основой неопластической трансформации клеток служит HPV-персистенция. Репликации вируса не происходит, и о его присутствии судят по наличию вирусных генов в базальном эпителии. Во многих случаях это не ведет к видимым патологическим изменениям. По разным данным, бессимптомная HPV-инфекция цервикального канала наблюдается у 10—40% женщин. Нередко повторное обследование через несколько месяцев дает отрицательный результат, отражая транзиторный характер носительства. Но латенция может быть и упорной, предрасполагая к осложнениям, главным из которых является канцерогенное (злокачественное) перерождение эпителия. У 15—30% женщин, позитивных по HPV, в течение двух лет развиваются предраковые изменения шейки матки; при отсутствии HPV-ДНК тот же показатель составляет 1—3%. Корреляция характерна для HPV-типов высокого риска, прежде всего 16 и 18. HPV-ДНК может присутствовать и в нормальных эпителиоцитах, прилегающих к папилломам. Это предрасполагает к рецидивам после удаления видимого очага опухоли.
Малигнизация не является внезапным событием. Как правило, это результат поэтапных, многолетних изменений эпителия — от незначительных до выраженных дисплазий. Инкубация составляет 10—20 лет, и лишь изредка сокращается до одного-двух лет. Предраковые состояния связаны с усилением пролиферации и подавлением дифференцировки эпителиоцитов базального слоя. Клетки обретают инвазивность, пенетрируя базальную мембрану. В отличие от доброкачественного онкогенеза (бородавки), когда избыточная пролиферация затрагивает все эпителиальные слои за исключением базальных клеток, при малигнизации она ограничена базальным клеточным слоем. Этому сопутствует подавление эпителиальной дифференцировки, что препятствует продуктивной репликации HPV. Экспрессия вирусного генома ограничивается транскрипцией наиболее ранних генов, прежде всего е6 и е7. Именно они кодируют главные вирусные онкопротеины, т.е. белки, ответственные за трансформацию клеток. С этой точки зрения Е6 и Е7 следует считать вирусными онкогенами.
Онкогенная стратегия канцерогенных папилломавирусов усиливается рядом механизмов, повышающих агрессивность опухолевого роста. Важной особенностью является интегративная вирогения, т.е. включение вирусного генома в состав клеточных хромосом. Интеграция, которая лишь изредка встречается при доброкачественных поражениях, типична для большинства инвазивных карцином и предраковых неоплазий1, существенно влияя на взаимоотношения вируса с клеткой. Во-первых, интегративная вирогения блокирует транскрипцию е2-гена, выключая синтез Е2-белка. Это снимает негативный контроль с е6- и е7-генов, создавая условия для гиперпродукции вирусных онкопротеинов. Во-вторых, HPV-ДНК может встраиваться по соседству с клеточными онкогенами, содействуя их активации. Наконец, интеграция содействует общей дестабилизации клеточного генома, предрасполагая к нерепарируемым мутациям. Накапливаясь, они ведут к селекции инвазивных клонов клеток. Об этом говорит цитогенетическая динамика HPV-индуцированной малигнизации: поликлональность первичных поражений сменяется олигоклональными неоплазиями и, наконец, моноклональными раковыми опухолями.
Важным признаком канцерогенных HPV является повышенное сродство их белков Е6 и Е7 к ингибиторам клеточной пролиферации и опухолевого роста, р53 и pRB. В сочетании с гипертранскрипцией е6- и е7-генов (из-за отсутствия е2-негативного контроля — см. выше) это ведет к срыву регуляторных механизмов в системах клеточного цикла и апоптоза, способствуя злокачественной трансформации клеток.
Онкогенез поддерживается внешними сигналами, действующими в кооперации с вирусными онкопротеинами. Как говорилось выше, HPV-трансформированные эпителиоциты обретают повышенную чувствительность к ростстимулирующим цитокинам и, соответственно, склонность к гиперпролиферации. Учитывая, что гены вирусных онкопротеинов (е6 и е7) у канцерогенных папилломавирусов гораздо агрессивнее, можно думать о значительно большем влиянии микроокружения (тех же цитокинов) на судьбу инфицированных ими клеток.
Присутствие канцерогенных HPV необходимо, но недостаточно для развития рака. Реализация их патогенности зависит от дополнительных факторов, которые могут усиливать или ослаблять трансформацию клеток. Малигнизации способствуют нарушения клеточного иммунитета (они возникают при ВИЧ-инфекции и фармакологической иммуносупрессии), длительное применение стероидных гормонов (оральные контрацептивы), герпетические инфекции, коинфицирование несколькими HPV-типами, многократная беременность, курение. Имеет значение и генетическая предрасположенность, влияющая на такие параметры инфекции, как чувствительность к заражению, способность к элиминации вируса, длительность инкубационного периода. Впрочем, по ряду позиций есть противоречия, и единственно надежным прогностическим критерием служит обнаружение канцерогенных разновидностей папилломавирусов, включая внутритиповые варианты, которые различаются по степени малигнизирующего эффекта, чувствительности к кофакторному сопровождению, географическому и этническому распространению.
Иммунитет. Большинство HPV-инфекций носят клинически латентный и преходящий характер. Лишь немногие получают развитие, вызывая опухоли, как правило, доброкачественной природы. Их естественная эволюция завершается спонтанной регрессией, для которой требуется разное время — от нескольких недель до несколько лет. Примерно половина кожных бородавок исчезает в течение двух лет. Около 90% HPV-индуцированных цервикальных изменений самоликвидируются на протяжении 12—36 мес. Это говорит о том, что иммунная система медленно, но в конечном счете эффективно справляется с инфекцией. Основу иммунной защиты составляют Т-лимфоциты. Судя по клеточному инфильтрату в зоне папиллом, главную роль играют CD4+-клетки и реакции замедленной гиперчувствительности. Динамика анти-HPV-антител не коррелирует с элиминацией вируса. Иммунный ответ не гарантирует от эндогенных рецидивов и от повторного заражения гетерологичными типами (и даже субтипами) HPV.
По мере малигнизации устойчивость клеток к иммунным реакциям возрастает, усиливая вероятность ракового перерождения. При слабой и умеренной дисплазиях цервикальные карциномы возникают в единичных случаях, тогда как не менее 12% агрессивных интраэпителиальных неоплазий трансформируются в инвазивные опухоли. Нередко это происходит на фоне анти-HPV-сдвигов (главным образом анти-Е6 и анти-Е7) в системах клеточного и гуморального иммунитета, не достигающих элиминационного эффекта. Одной из причин может быть лимитированная экспрессия вирусных генов и ее снижение по мере развития диспластического процесса.
Диагностика и лечение
HPV-инфекция легко диагностируется визуально — на основе клинических и патогистологических признаков. Коммерчески доступны антитела для иммуноцитохимического определения общего (родоспецифического) антигена папилломавирусов, но их чувствительность недостаточна для надежного диагноза. Серологическая диагностика (определение анти-HPV-антител) тоже малочувствительна и вряд ли получит широкое применение. Выделение вирусов — слишком сложная процедура для рядовых лабораторий.
Чаще используется молекулярно-генетический анализ (выявление HPV-ДНК). Это выглядит целесообразным для прогностического обнаружения канцерогенных HPV при эпителиальных дисплазиях шейки матки.
Лечение основано на элиминации HPV-папиллом химическими и физическими методами. В первом случае применяется обработка кислотами, формальдегидом, глютеральдегидом, антиметаболитами. Для физической эрадикации используют механическое удаление, криотерапию (обработку жидким азотом), лазерную и электрохирургию. Терапевтический эффект оказывают местные аппликации интерферонов и их индукторов (в виде мазей). Однако возможны рецидивы, связанные с персистенцией вируса в сопредельных тканях.
Признание причинной связи между HPV и злокачественной трансформацией клеток, а также естественная элиминация папилломавирусов на этапе доклинической латенции, доброкачественных и предраковых неоплазий позволяют надеяться на возможность разработки специфической профилактики и терапии HPV-ассоциированных патологий, прежде всего цервикального рака. В качестве профилактических вакцин планируется использовать вирусоподобные частицы, полученные из капсидных (L1 и L2) белков канцерогенных папилломавирусов. Их превентивная активность связана с антителами, нейтрализующими вирионы.
Для лечения разрабатываются препараты на основе Е6/Е7-онкопротеинов. Это логично, если учесть, что они необходимы для индукции и стабилизации злокачественного фенотипа клеток и к тому же являются единственными HPV-белками, экспрессируемыми в клетках цервикальной карциномы. Одна из проблем связана с неоднородностью папилломавирусов. Это потребует поливалентных вакцин с учетом антигенной специфики канцерогенных папилломавирусов. Главное внимание сконцентрировано на двух наиболее опасных HPV-типах — 16 и 18. На основе их капсидных белков (прежде всего L1) получены вирусоподобные частицы (капсиды без ДНК), которые проходят испытания в качестве профилактических и лечебных вакцин.
Ротавирусы
У всякого чина своя причина.
- Вирусная природа острого гастроэнтерита.
- Ротавирусы — лидер острой диареи у самых маленьких детей.
- Этимология терминов Rotavirus и Reoviridae.
- Вирион: группы, подгруппы, типы.
- Репродукция ротавирусов.
- Особенности патогенеза и иммунитета.
- Диагностика и неудачи специфической профилактики.
Hаряду с острыми инфекциями верхних дыхательных путей инфекционная диарея принадлежит к числу наиболее распространенных заболеваний человека, прежде всего детей. Она называется острым гастроэнтеритом, объединяя широкий круг инфекций вирусной, бактериальной и протозойной природы. Картина может варьировать от гастроинтестинальных симптомов без воспалительного поражения энтероцитов до воспаления с обильной, но обычно умеренной диареей. Присоединяются разнообразные симптомы, включая абдоминальные боли и спазмы, слабость, отсутствие аппетита, миалгию, тошноту, которые появляются отдельно или вместе с диареей и рвотой. В тяжелых случаях (как правило, среди младенцев и детей до двух лет) заболевание приводит к потере жидкости, опасной для жизни больного. Ежегодно регистрируется 1—2 млрд. диарейных заболеваний, более 60% заболевших составляют дети; умирает до 3,5 млн. человек. Это большая проблема для всего мира, особенно для развивающихся стран. В развитых странах острый гастроэнтерит занимает второе место среди общей клинической патологии, уступая лишь инфекционным заболеваниям верхних дыхательных путей.
Главная роль в этиологии острого гастроэнтерита принадлежит вирусам. Утверждение этой позиции началось в 1940-х гг., но окончательно состоялось лишь в начале 1970-х гг., когда в фекалиях детей с гастроэнтеритом были обнаружены вирусы Норуолк (Norwalk) и ротавирусы и доказано их отношение к данному заболеванию. В настоящее время известно четыре семейства вирусов, которые способны вызывать острый гастроэнтерит у человека — ротавирусы (Reoviridae), калицивирусы, включая Норуолк (Caliciviridae), аденовирусы (Adenoviridae, типы 31, 40, 41 — см. «Аденовирусы») и астровирусы (Astroviridae). Подозревается значение и других вирусов (например, торовирусов, коронавирусов, энтеровируса 22 и др.). Их роль в происхождении гастроэнтерита не доказана, хотя вполне вероятно, что будут открыты новые вирусные агенты, принимающие участие в интестинальной патологии, — этиология 40—50% диарей остается нераскрытой. Основными возбудителями острого гастроэнтерита являются ротавирусы.
Ротавирусы получили свое название из-за того, что под электронным микроскопом напоминают колесо с короткими спицами и хорошо развитым ободом (лат. rota — колесо; рис. 1).

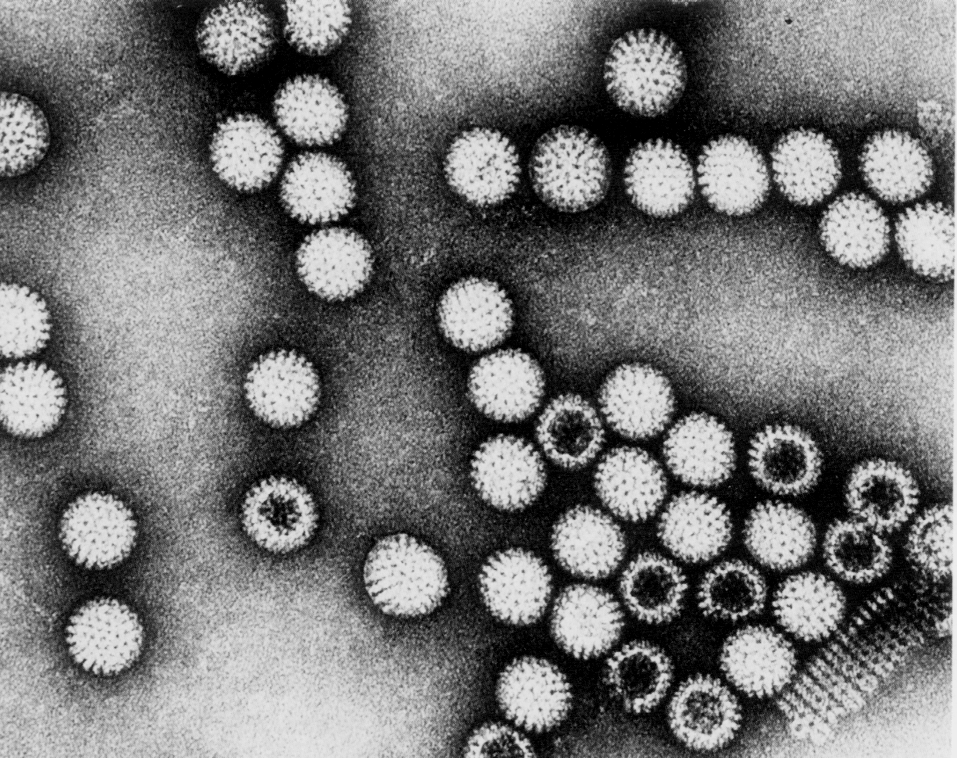
Позже оказалось, что по своему строению и жизненному циклу ротавирусы похожи на реовирусы. В 1978 г. они были выделены в специальный род, Rotavirus, семейства Reoviridae. Это семейство включает девять родов. Четыре из них (Orthoreovirus, Rotavirus, Coltivirus, Orbivirus) способны инфицировать человека, но заболевания вызывают почти исключительно ротавирусы. Представители четырех других родов инфицируют растения и насекомых; один род связан с поражением рыб.
У термина «реовирусы» интересная этимология. Реовирусы были обнаружены в респираторном и кишечном трактах человека и животных, но их значение оставалось неизвестным. Поэтому они были названы реовирусами от слов «респираторный», «энтеральный» и «orphan» (англ. orphan, сирота), т.е. вирусы с неустановленной ролью в патологии.
Вирион и репродукция ротавирусов
Вирион ротавирусов имеет диаметр 70—75 нм и состоит из трех структурных элементов — сердцевины (кора), наружного и внутреннего капсидов с икосаэдрической симметрией. Капсомеры внутреннего капсида выглядят как радиальные спицы, образуя подобие колеса, где наружный капсид служит ободом. Кор содержит геномную РНК и внутренние белки — VP2 (главный структурный компонент внутренней мембраны), VP1 и VP3 (белки, отвечающие за РНК-зависимую РНК-полимеразную активность и кэппинг вирусной мРНК). Как и у других реовирусов, геном ротавирусов представлен 11 сегментами минуснегативной двунитчатой РНК с общей массой более 18000 пар оснований. Каждый из сегментов кодирует единственный вирусный белок, т.е. является моноцистронной матрицей. Исключением служит сегмент VP7 — бицистронный ген, в котором записано два гликопротеина с двух инициирующих кодонов в одной рамке считывания.
Внутренний и наружный капсиды содержат три белка — VP6, VP7 и VP4. Это главные антигены, которые используются в классификации ротавирусов и служат началом для взаимодействия вириона с клетками-мишенями. VP4 и VP7 называются нейтрализующими антигенами, так как анти-VP4- и анти-VP7-антитела блокируют вирусную инфекцию in vitro. При обработке трипсином VP4 расщепляется на два пептида, VP5* и VP8*, что ведет к активации инфекционных свойств вириона и возможности культивирования ротавирусов in vitro.
Рецепция вириона происходит достаточно сложно. Сначала в реакцию включается VP8 (продукт трипсинного расщепления VP4), который связывается с сиаловой кислотой или ганглиозидом GM1a инфицируемой клетки. Далее происходит взаимодействие VP4 и VP7 с a2b1— и a4b1-интегринами. Затем в реакцию включаются другие клеточные гликопротеины, гликолипиды и холестерол, которые формируют зону восприятия вируса. Вирион проникает путем эндоцитоза, используя VP4 (VP5) и VP7 для пробуравливания плазматической мембраны клетки.
Процесс раздевания вируса продолжается внутри клетки. Низкая концентрация Са2+ в рецептосоме ведет к дезинтеграции наружного капсида, обеспечивая выход в цитоплазму однокапсидной вирусной частицы. В ее составе происходит активация РНК-зависимой РНК-полимеразы и синтез плюс-нитей РНК, которые кэпируются и выделяются через многочисленные поры однокапсидного вириона. Готовые плюс-РНК (мРНК) идут на образование структурных и неструктурных вирусных белков. Неструктурные белки (NSP1—NSP6) участвуют в самосборке однокапсидной субвирусной частицы, синтезе минус-нитей РНК, ее почковании через гранулярную эндоплазматическую сеть и, наконец, освобождении полных двухкапсидных вирионов в результате клеточного лизиса. Частица временно обретает псевдооболочку, которая быстро замещается белками наружного капсида, т.е. VP4 и VP7. Весь процесс занимает 6 ч, достигая максимума через 15—24 ч.
VP6 представляет собой белок внутреннего капсида, связывая в единое целое коровый белок VP2 и белки наружного капсида, VP4 и VP7. Он несет групповые эпитопы (A—G), хотя все ротавирусы связаны антигенным родством. В медицинской микробиологии наибольшее значение имеет группа А, которая отвечает почти за 100% детского ротавирусного гастроэнтерита. Ротавирусы В известны как зоонозные возбудители диарей у взрослых и детей (источник инфекции — крысы), эндемичны для Китая. Были сообщения о вспышках ротавирусной инфекции группы С. Возможно, заражение произошло от свиней, которые выделяют эти ротавирусы.
Вирусы группы А делятся на подгруппы и множество серотипов. По особенностям VP6 выделяют по меньшей мере четыре подгруппы, из которых для человека важны подгруппы I и II. Но большее значение имеет деление на серотипы, дающее возможность судить об основных протективных антигенах вируса, задействованных в патологии. Это важно для суждения об эпидемиологии заболеваний и подбора иммунологических препаратов с целью их профилактики и лечения. Сегодня известно 14 VP7-серотипов (G-серотипы, от англ. glycoprotein) и 20 VP4-типов (Р-типы, от англ. protease-sensitive protein). Не все Р-типы подтверждены как серотипы, поэтому применяют два понятия — Р-серотипы и Р-генотипы. Между ними имеется ряд важных, но не абсолютных совпадений. Например, наиболее распространенные Р-серотипы 1А и 1В соответствуют генотипам 8 и 4, в то время как генотипы 2 и 3 могут попеременно встречаться с серотипом 5В. Поэтому предложена двойная система классификации ротавирусов по VP4, которая учитывает серотип и генотип VP4. При ее использовании G-Р-серотип дополняется в скобках Р-генотипом, например, G1Р1А [8]. Часто ограничиваются обозначением Р-генотипа, G1P8. Человека инфицирует ограниченное число G-P-вариантов. Доминирующими являются G1-4-серотипы и Р4-, Р6-, Р8-генотипы (их различают по полимеразной цепной реакции).
Так как VP7 и VP4 кодируются разными РНК-сегментами, они могут независимо перемещаться, давая множество G-P-комбинаций в процессе пересортировки генов in vitro и in vivo.
К примеру, G3 может нести детерминанты по крайней мере пяти Р-типов (Р2, Р3, Р8, Р12, Р16), а Р8-тип — четырех различных G-серотипов (G1, G3, G4, G9). Некоторые G-P-типы определяются только у человека (например, G9, G12, P4, P6, P8) или у животных (G11, G13, P3, P11). Известны комбинации, которые наблюдались как у людей, так и у животных, что допускает вероятность зоонозной трансмиссии.
Для дифференцировки различных изолятов предложено использовать электрофоретический профиль геномных РНК-сегментов в полиакриламидном геле — так называемый электроферотип (напомним, что геном ротавирусов представлен 11 сегментами РНК). В многочисленных исследованиях продемонстрирована высокая геномная гетерогенность ретровирусов, которая оказалась стабильным признаком штамма, полезным для эпидемиологического анализа. Кроме длинных и коротких электроферотипов (по сравнительной скорости миграции РНК-сегментов 10 и 11) наблюдаются особенности передвижения отдельных компонентов (рис. 2). Детали электроферотипов не совпадают с серотипами, оставляя за последними приоритет в разработке методов контроля над инфекцией.

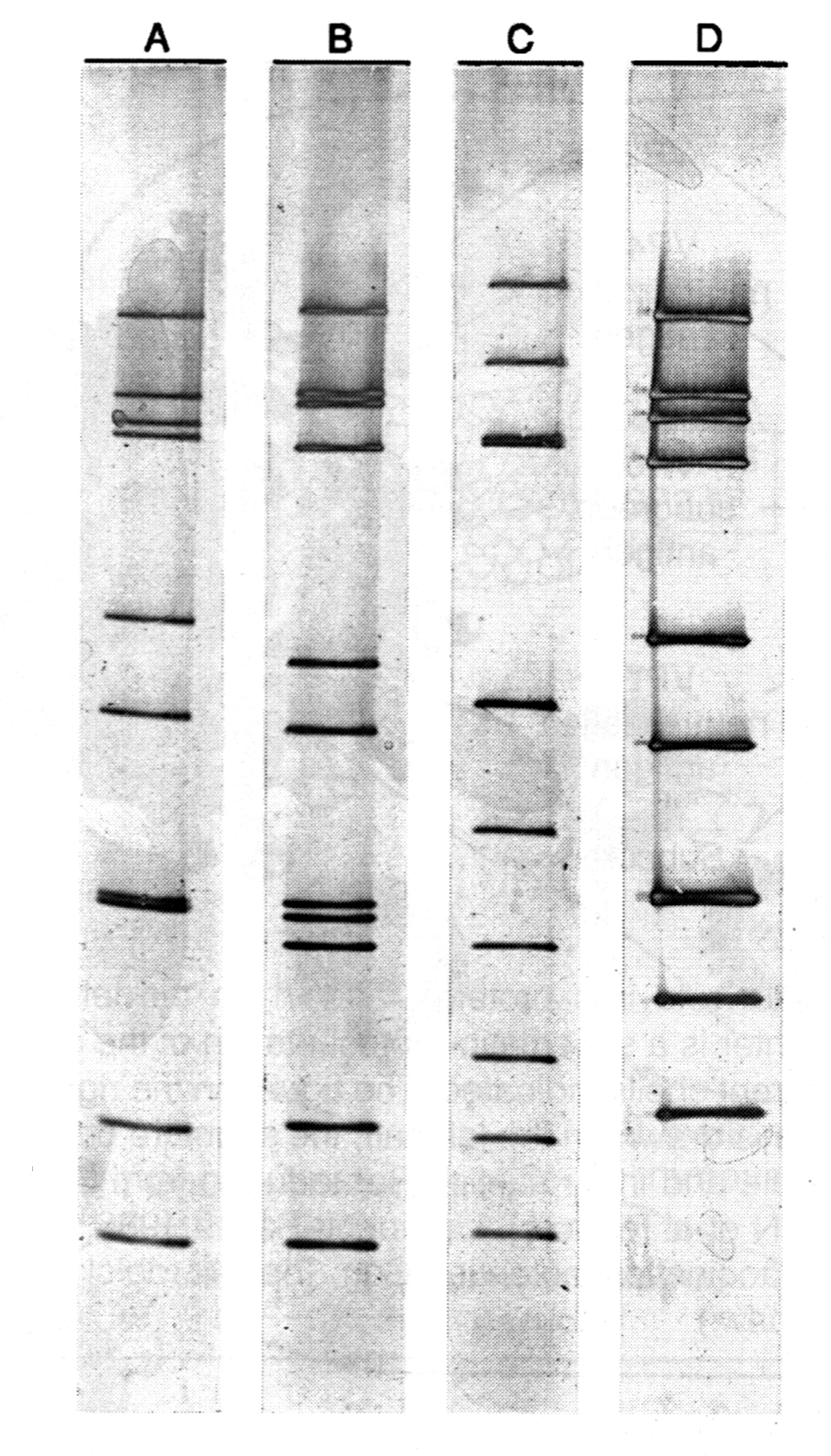
Эпидемиология
Ротавирусы являются ведущим фактором гастроэнтерита у многих видов молодых животных, включая человека. Они распространены во всем мире, отвечая примерно за 50% случаев острого гастроэнтерита у госпитализируемых детей. Судя по антителам, к трем годам более 90% детей уже были в контакте с главными серотипами вируса. Источником заражения могут быть старшие дети или взрослые с субклинической инфекцией. Впрочем, выделение огромных количеств вируса в остром периоде болезни (1011/г фекалий) и достаточность для инфицирования всего нескольких вирионов создает возможность заражения от больного ребенка. Вероятность инфекции повышается благодаря устойчивости вируса во внешней среде. Ротавирусы высокорезистентны к большинству общеупотребляемых дезинфектантов.
В развивающихся странах ежегодно ротавирусы отвечают за 140 млн. случаев и 800 000 смертей от острой диареи. В развитых странах, например в США, регистрируется около 3 млн. детских ротавирусных гастроэнтеритов в год, 150 из них заканчиваются летально. В России во время осенне-зимнего подъема ротавирусы вызывают 60—90% острых кишечных инфекций у детей до трех лет. Клинически картина ротавирусного гастроэнтерита не отличается от кишечных инфекций другой этиологии. Болеют главным образом дети от 6 мес до 2 лет, иногда пожилые люди после 60 лет. Взрослые обычно являются бессимптомными вирусоносителями. Убиквитарность ротавирусов определяет почти поголовное инфицирование детского населения. Взрослые сохраняют приобретенный уровень иммунитета, что позволяет думать о субклинических реинфекциях, поддерживающих иммунный статус.
Клиника, патогенез и иммунитет
Ротавирусная инфекция ограничивается наиболее диффенцированными клетками апикальных отделов ворсинок тонкой кишки. Заражение происходит благодаря фекально-оральному механизму, который реализуется различными, но принятыми для кишечных инфекций путями — контактно-бытовым, пищевым, водным. Инкубационный период не превышает 48 ч. Патологические изменения кишечника включают укорочение и атрофию ворсинок, реактивную гиперплазию крипт (кубические незрелые клетки крипт замещают погибающие клетки ворсинок), мононуклеарную инфильтрацию собственной пластинки и расширение цистерн эндоплазматической сети. Но между гистопатологией и симптомами не обнаружено прямой корреляции, что заставляет искать дополнительные причины ротавирусной диареи. Одним из механизмов может быть то, что ротавирусы, повреждая энтероциты, индуцируют мальабсорбцию углеводов, которая способствует осмотической диарее. К этому присоединяется диарея секреторного типа, связанная с гиперплазией функционально незрелых криптических клеток. Другим объяснением является повышение мукозальной проницаемости за счет структурного и функционального изменения межклеточных контактов (без нарушения клеток). Эту роль выполняет NSP4 — первый и пока единственный из вирусных энтеротоксинов.
Еще одна гипотеза постулирована недавно. Она говорит о том, что ротавирусы усиливают интестинальную секрецию, активируя нервную систему кишечной стенки. Это подтверждено клиническими наблюдениями с использованием ингибиторов энтеральной нервной системы. Имеет значение и чувствительность капсидных антигенов (VP4) к трипсину, которая ведет к повышению инфекционной активности вируса.
Начало симптомов — острое, период инкубации составляет 1—2 дня. У большинства детей заболевание проявляется в виде диареи и рвоты, которые продолжаются 2—6 дней. Этому нередко сопутствуют симптомы поражения дыхательного тракта (ринит, ринофарингит, фарингит), хотя их природа остается неясной (ротавирусы не размножаются в респираторном эпителии). Клинические признаки варьируют от мягких симптомов до очень тяжелых, что частично зависит от дозы инфекции и природы ротавирусного штамма. Особенно тяжело заболевание протекает на фоне иммунодефицита и у детей, ослабленных голоданием.
Ротавирусная инфекция сопровождается местным (интестинальным) и общим иммунным ответом. Полагают, что в течение первых трех месяцев жизни материнские IgG-антитела защищают новорожденного от клинически значимой инфекции, хотя по другой версии, серьезное значение имеет защита, создаваемая в кишечнике ребенка лактатгерином — гликопротеином человеческого молока.
Главная протективная роль принадлежит мукозальным антителам. Однако интестинальные IgA-антитела действуют недолго, а IgA-память отстает от вируса, который, вызывая инфекцию с коротким инкубационным периодом, успевает проникнуть и поразить энтероциты. Это объясняет широкое реинфицирование детей, чему, безусловно, способствует антигенная неоднородность и изменчивость ротавирусов. И все-таки после трех лет постоянных контактов с вирусом организм обретает к нему устойчивость. Заражение обычно ведет к бессимптомной инфекции, которая поддерживает циркуляцию ротавирусов у человека. Попытки найти соответствие между резистентностью и нейтрализующими VP4 и VP7 антителами не привели к однозначным результатам. Скорее всего это связано с тем, что в защите от вируса участвуют интестинальные и сывороточные антитела к другим вирусным белкам, в частности VP6 и NSP4. В настоящее время отсутствует точный и надежный маркер, по которому можно было бы судить об эффективности иммунитета против ротавирусов.
Профилактика, диагностика и лечение
Строгая изоляция ротавирусных больных не является обязательной мерой для предотвращения ротавирусной инфекции. Ротавирусы высокорезистентны ко многим обычно используемым дезинфектантам, так что едва ли не единственной надежной мерой борьбы с заражением является тщательное мытье рук. Впрочем, наблюдение о том, что число ротавирусных инфекций примерно одинаково в развитых и развивающихся странах (в первом случае процент фатальных исходов гораздо ниже), говорит о том, что неспецифические процедуры не отменяют необходимости в вакцинации.
К сожалению, вакцина на основе обезьяньего ротавируса G3 и трех человеческих монореассортантов (G1, G2, G4) была забракована из-за редких осложнений, связанных с инвагинацией кишечника. Новые вакцины предполагают использование аттенуированных и рекомбинантных штаммов человека, применение рекомбинантных и синтетических ротавирусных антигенов, ДНК-вакцин. Некоторые из них (например, 89-11-вакцина из аттенуированного человеческого штамма G1P8) проходят интенсивные испытания. В тяжелых случаях, чтобы избежать дегидратации, назначают поддерживающую оральную (реже внутривенную) регидратационную терапию. ВОЗ для оральной регидратации рекомендует следующий раствор:
- хлорид натрия — 3,5 г;
- хлорид калия — 1,5 г;
- цитрат натрия — 2,9 г;
- глюкоза — 20,0 г.
Все это растворяется в 1 л чистой питьевой воды.
В остальном полагаются на здоровое питание и на то, что ротавирусная инфекция обычно самоизлечивается через 1—2 нед.
На пике болезни вирус выделяется в таком количестве, что его легко выявить в фекалиях больного. Применяют агглютинационные тесты на основе частиц латекса, покрытых ротавирусными антителами, и различные варианты иммуноэнзимного анализа (РЭМА). Чувствительность латексной агглютинации вполне достаточна для раннего периода инфекции; позже требуется подтверждение результата в РЭМА. Определение групповых, подгрупповых антигенов, а также G-P-серотипов проводят в РЭМА при помощи моноклональных антител. Недавно для этого стали применять обратно-транскриптазную полимеразную цепную реакцию с гено- и типоспецифическими праймерами. Большинство человеческих штаммов группы А принадлежат к подгруппе II, серотипам G1P8, G3P8 или G4P8, или к подгруппе I, серотипу G2P4.
Возможно применение электронной микроскопии. Метод широко использовался до внедрения в практику латексной агглютинации и РЭМА, но теперь почти не применяется из-за сложности с оборудованием, более низкой чувствительности и трудоемкости. То же самое можно сказать об идентификации вируса по электрофорезу РНК, выделенной из экстракта испражнений.
Для определения титра антител у больных детей и субклинически инфицированных ротавирусами можно использовать различные серологические тесты, прежде всего на основе РЭМА.
Вирус бешенства
Та же птица, да песни другие.
- История болезни и возбудителя.
- Современная классификация.
- Эпидемиология и эволюция рабической инфекции.
- Вирион и репликация.
- Патогенез: от персистенции до поражения ЦНС.
- Клиника и профилактика бешенства.
Бешенство у собак известно с глубокой древности. Например, еще в «Илиаде» греки называли троянца Гектора бешеной собакой. Некоторые аспекты эпидемиологии были упомянуты в трудах Аристотеля, Демокрита (322 г. до н.э.) и др. Первое клиническое описание заболевания у человека принадлежит Цельсу (I в. н.э.), который назвал ее водобоязнью (hydrophobia) и рекомендовал прижигать укушенные раны. В 1804 г. Цинке экспериментально доказал возможность передачи заболевания со слюной больного животного. Распространение возбудителя в зараженном организме по нервным стволам выявлено Бабешем (1887 г.). Вирусная этиология этого заболевания была установлена в 1903 г. Ремленже.
Благодаря работам Пастера выявлена локализация возбудителя в организме (ЦНС), заложены основы специфической профилактики бешенства (1885 г.). Путем многочисленных экспериментов на животных Пастеру и сотрудникам удалось получить так называемый фиксированный вирус, который использовался в дальнейшем для приготовления вакцин. Для этого уличный вирус бешенства пассировался интрацеребрально на кроликах. По мере пассирования длительный инкубационный период, характерный для инфекции, вызванной уличным вирусом, постепенно сокращался, пока не стал составлять 5 дней. Этот вирус, названный фиксированным (virus fixe), при интрацеребральном введении вызывал 100% гибель кроликов. Вместе с тем для собак и человека он частично потерял вирулентность, и после дополнительной обработки (высушивание мозга кролика над едким натром, способствующее полной инактивации вируса) фиксированный вирус был использован в качестве вакцины для лечебно-профилактических прививок. Первая пастеровская станция в России (г. Одесса), где готовили антирабическую вакцину, была создана в 1886 г. И.И. Мечниковым и Н.Ф. Гамалея.
Возбудитель бешенства Neuroryctes rabiei относится к семейству Rhabdoviridae (греч. rhabdos — прут, палочка) рода Lyssavirus (греч. lyssa — бешенство). Рабдовирусы представляют собой наиболее обширную и разнообразную группу среди семейств вирусов, имеющих минус-РНК. Она обширна не только по числу входящих в нее вирусов, но и по количеству поражаемых хозяев: растения, простейшие, насекомые, млекопитающие, птицы, рыбы. Несмотря на такой разброс экологических ниш, оказалось, что все эти вирусы имеют много общего: величина генома, строение вириона, механизм репликации. Главным критерием их классификации служит морфология: все рабдовирусы вытянуты в длину и имеют форму пули или длинной палочки с закругленными концами, что отражено в названии семейства.
Впервые термин «рабдовирусы» был применен на заседании Международного комитета по таксономии в 1971 г. для обозначения семейства вирусов, имеющих пуле- или палочковидную форму, спиральный нуклеокапсид, окруженный оболочкой с поверхностными выступами (шипами). В семейство рабдовирусов было включено около 60 вирусов с разными биологическими свойствами. В 1975 г. семейство разделили на три рода: Vesiculovirus (вирус везикулярного стоматита), Lyssavirus (вирусы бешенства) и рабдовирусы растений.
Способность рабдовирусов размножаться в клетках членистоногих и млекопитающих позволила рассматривать некоторые из них в качестве возбудителей арбовирусных инфекций. Род Lissavirus включает многочисленные вирусы, сходные в антигенном отношении, но различающиеся по спектру хозяев, вирулентности, инфекционному титру, адаптационным свойствам, способности к образованию цитоплазматических включений (тельца Негри). Комитет экспертов ВОЗ по бешенству предложил классифицировать их следующим образом:
- Серотип 1 — прототипный штамм CVS 24, включает большинство уличных штаммов из различных частей света, а также выделенные от грызунов в Центральной Европе.
- Серотип 2 — прототипный штамм Lagos Bat, выделенный из мозга летучих мышей в Нигерии, Центрально-Африканской республике, Гвинее и от кошки в Зимбабве.
- Серотип 3 — прототипный штамм Mokola, изолированный от землеройки в Нигерии, от человека в Камеруне, от диких и домашних животных в Зимбабве.
- Серотип 4 — прототип Duvenhage, изолирован от человека, укушенного летучей мышью в ЮАР, а также от летучих мышей в Южной Африке и Центральной Европе.
- Серотип 5 — Lyssavirus, выделенный от европейских мышей и от укушенных ими людей в Финляндии и СССР.
Подобно буньявирусам, рабдовирусы возникли, по-видимому, как вирусы заболеваний, передаваемых членистоногими. Некоторые представители данного семейства еще сохранили способность передаваться через насекомых, в то время как вирус бешенства эту способность потерял.
Эпидемиология
Бешенство — типичный зооноз, из группы инфекционных болезней, возбудители которых передаются при укусе и/или ослюнении поврежденных кожных покровов (реже — слизистых оболочек). Таким образом, бешенство можно отнести к раневым инфекциям, несмотря на экспериментальные данные, которые свидетельствуют о возможности осуществления нераневых путей передачи (перорального, аэрогенного). В естественных условиях вирус попадает в организм реципиента в основном при прямом контакте. Восприимчивость к бешенству всеобщая, заболевание встречается во все времена года. В эпизоотологии инфекции различают природный тип бешенства и антропургический (городской). Основным источником и резервуаром первого типа являются дикие плотоядные животные (лисица, песец, волк, шакал, енотовидная собака, енот, мангуст), а в странах Америки — летучие мыши, среди которых возможен аспирационный механизм передачи (в основном летучие мыши-вампиры). При втором типе бешенство поддерживается бродячими собаками, кошками, а также сельскохозяйственными животными.
Основным источником рабической инфекции остается собака не только в Азии, но также в Африке и некоторых странах Латинской Америки. В нашей стране описаны случаи заражения бешенством людей после укусов летучих насекомоядных мышей. Лиссаподобный вирус от насекомоядной летучей мыши также был выделен в английском порту Ньюхавен в 1996 г.
В европейских странах (в том числе в России) после 40-х гг. прошлого века отмечена эволюция рабической инфекции — формирование крупных, стойких, постоянно мигрирующих очагов лисьего бешенства. В течение 40—50 гг. они оккупировали огромную территорию Европы — от Польши до Франции, Бельгии, Нидерландов, Швейцарии, мигрируя в среднем со скоростью 40 км в год. Возможно, среди лисиц циркулируют генетически измененные штаммы дикого вируса. Замечено, что бешенству у лисиц предшествует нашествие грызунов. Среди мышей, крыс, птиц и собак известны случаи абортивной и бессимптомной форм бешенства, что подтверждается и экспериментально. В нашей стране первые очаги лисьего бешенства были замечены в Тамчикском заповеднике Астраханской области, Московской и Калининградской областях.
В нашей стране очаги лисьего бешенства занимают огромную территорию от Калининградской области до Дальнего Востока, свободны лишь ограниченные территории: Ярославская, Костромская, Ивановская, Томская и другие области. На Крайнем Севере очаги арктического бешенства поддерживаются главным образом песцами. В последние годы обстановка по бешенству среди животных на территории Российской Федерации обострилась, в связи с этим значительно возрастает угроза распространения этой инфекции среди населения. Ситуация усугубляется ростом численности безнадзорных животных, нарушением правил содержания домашних животных и ввоза из-за рубежа. Наиболее стойкие природные очаги сохраняются в Центральном, Центрально-Черноземном, Северо-Кавказском, Поволжском, Уральском, Дальневосточном районах страны. В этих же районах регистрируются случаи заболевания людей этой инфекцией. Число лиц, получивших повреждения от животных в 90-е гг., по сравнению с 80-ми увеличилось более чем в два раза.
В Российской Федерации ежегодно регистрируется от 5 до 20 случаев бешенства среди людей. За 1996 г. погибло 16 человек (Астраханская, Московская, Курская, Самарская области и др.). В 1997—1999 гг. в Москве зарегистрировано 3 случая смерти людей от бешенства.
Инфекция не зарегистрирована в Австралии, Новой Зеландии, на Гавайских островах, Ямайке, Кипре, Мальте, а также в Норвегии, Швеции, Финляндии, Испании и Португалии.
Вирион и репликация
Строение и внешний вид вириона возбудителя бешенства типичны для рабдовирусов (см. рисунок). Это частицы пулевидной формы, их размер 80—180 нм, имеют спиральный тип симметрии нуклеокапсида, относятся к оболочечным вирусам. Геном представлен одноцепочечной РНК (мол. масса 4,6·106 Да), которая упакована в комплексе с белком в виде спирали и окружена внешней оболочкой. На поверхности вирионов имеется множество мелких шипов. РНК, как у всех рабдовирусов, обладает негативной полярностью и имеет необходимую для синтеза мРНК РНК-зависимую РНК-полимеразу.

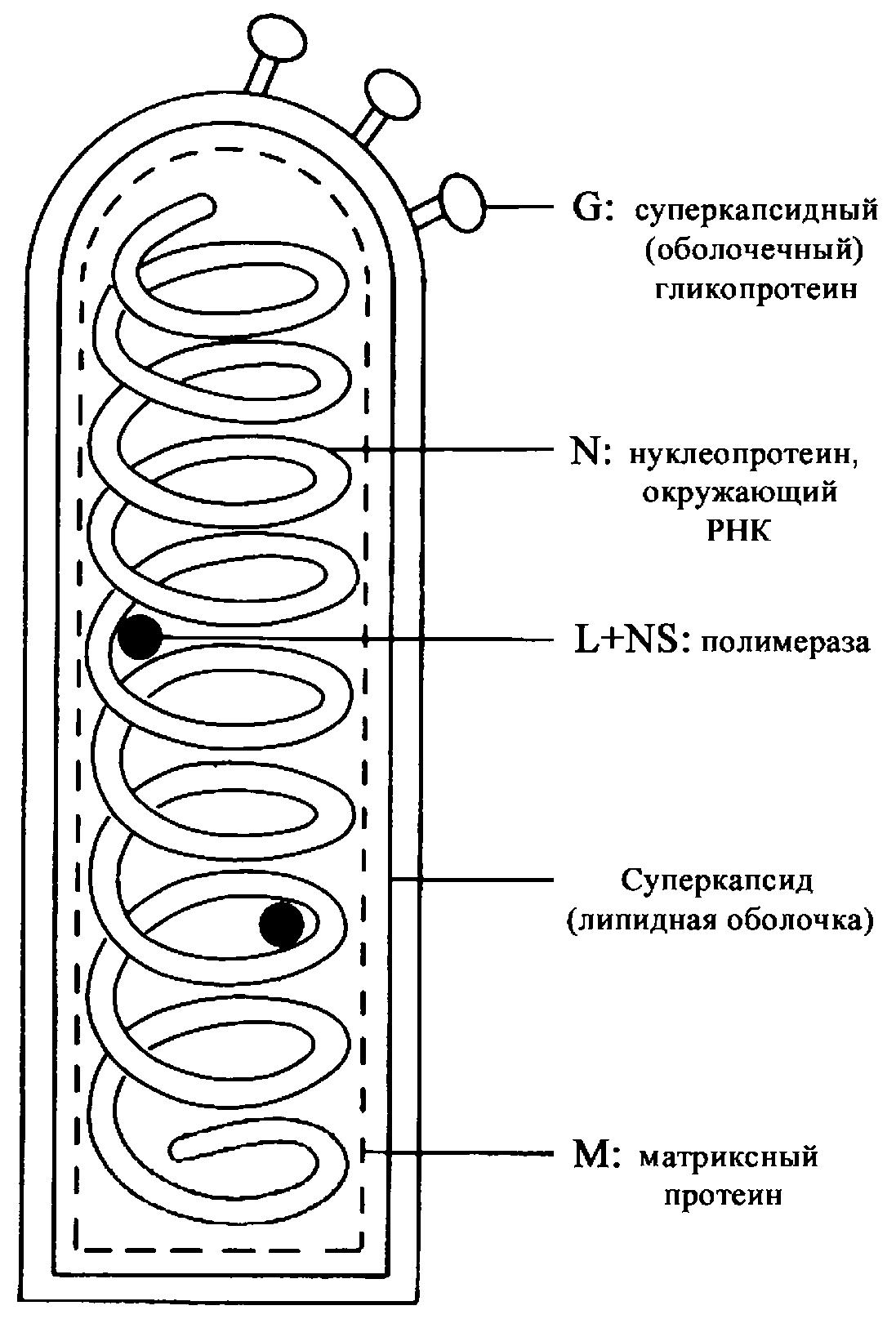
Вирион располагает пятью разновидностями белков, которые делятся на внутренние и поверхностные. К внутренним относятся белки N, M, L, NS. Белок нуклеокапсида (N, от англ. nucleocapsid) непосредственно связан с РНК и серологически идентичен у всех рабдовирусов. Одной из его функций является формирование рибонуклеопротеина, в составе которого РНК обретает устойчивость к клеточной РНК-азе. Он индуцирует образование комплементсвязывающих антител.
С нуклеокапсидом связано еще два белка сердцевины: L (от англ. large — большой) и NS (от англ. non structural — неструктурный), которые являются компонентами транскриптазы и в совокупности функционируют как полимераза. Белок L обеспечивает инициацию синтеза РНК, а NS-белок — их элонгацию.
Матриксный белок М (от англ. matrix) составляет внутренний слой липопротеиновой оболочки (суперкапсида) вириона, связывая его с сердцевиной. Его функцией является стягивание звеньев рибонуклеопротеина в цилиндр со спиральной симметрией. Белок М принимает участие в завершающей стадии морфогенеза вируса в зараженной клетке.
Поверхностный белок G (от англ. glycoprotein) представлен гликопротеином, входит в состав оболочки (суперкапсида) и структурно оформлен в виде колб диаметром 4,5—5,5 нм и длиной ножки 2 нм. Качественный состав белковой части молекулы варьирует в зависимости от вирулентности вируса. Например, в более вирулентном варианте аргинин замещается глутамином и изолейцином. Играет ведущую роль в процессах адсорбции на поверхности чувствительных клеток. G-белок является протективным антигеном, т.е. индуцирует образование вируснейтрализующих антител, которые блокируют адгезию вируса. Он также обладает гемагглютинирующими свойствами.
Вирион, прикрепляясь белком G к рецепторам чувствительной клетки, проникает в нее путем эндоцитоза, а не вследствие слияния суперкапсида с ее мембраной (у рабдовирусов нет белка, ответственного за этот процесс). Внутри клетки вирус обнаруживается в эндосомах, где при кислых значениях рН меняется конформация его G-белка, делая его чувствительным к клеточным протеазам. В результате ограниченного протеолиза обнажаются пептиды, вызывающие слияние суперкапсида вируса и эндосомальной мембраны. Благодаря этому нуклеокапсид освобождается и выходит в цитоплазму.
После раздевания начинается транскрипция. В эукариотических клетках нет РНК-зависимой РНК-полимеразы, поэтому для экспрессии своего генома рабдовирусы используют собственную полимеразу. Матрицей является РНК, прочно связанная с N-белком. Геном транскрибируется с образованием пяти моноцистронных мРНК. Это приводит к появлению мРНК, пригодных для трансляции, и в то же время позволяет регулировать количество отдельных мРНК. Каждый вирусный белок проходит свой путь созревания.
Репликация осуществляется в цитоплазме в две стадии. Минус-цепь в составе рибонуклеокапсида служит матрицей для синтеза комплементарной РНК, совпадающей по размеру с геномной. N-белок связывается с новосинтезированной РНК, образуя рибонуклеокапсид, содержащий плюс-геномную РНК. Затем процесс повторяется, и на плюс-РНК, как на матрице, образуются новые минус-цепи. В процесс репликации вовлечены N-, NS- и L-белки. Поскольку матрицы для транскрипции и для синтеза плюс-цепей в ходе репликации одинаковы, не удивительно, что эти процессы контролируются одними и теми же белками. Сердцевина вируса, состоящая из РНК, белков N, NS и L, собирается как отдельная структура в цитоплазме вместе с репликацией вирусной РНК и синтезом структурных (вирионных) белков.
Почкование зрелых вирионов — сложный процесс, для которого необходимы ассоциированный с мембраной G-белок, М-белок и рибонуклеопротеиновые частицы. Все они собираются на определенном участке мембраны. Ключевую роль в сборке и почковании играет М-белок. Он связывает рибонуклеопротеин и оболочку, кроме того, способен вызывать сворачивание удлиненных рибонуклеопротеиновых частиц с образованием той уплотненной конфигурации, которая присутствует в вирионах.
В системах in vitro и in vivo у вируса бешенства происходит формирование двух типов частиц: В (от англ. bullet — пуля) — зрелые полные вирионы и Т ( от англ. trunk — ствол) — дефектные интерферирующие частицы. Последние не обладают инфекционностью, имеют размер 60—80 нм и содержат приблизительно половину вирусного генома. Дефектные частицы размножаются только в присутствии полного вируса (В-частиц). Интерференция с В-частицами происходит на стадии репликации вирусной РНК.
Патогенез
Заражение происходит при укусе и/или ослюнении раны больным бешенством животным. Некоторое время (от нескольких часов до 6 дней) вирус находится в месте внедрения. Здесь он концентрируется в мионевральных синапсах и первичное размножение происходит в миоцитах или в эпителиальных клетках базального слоя эпидермиса. Далее вирус проникает в сенсорные и двигательные нервные окончания путем связывания с ацетилхолиновыми рецепторами нервно-мышечных синапсов. В этом месте нервные окончания не имеют миелиновой оболочки, что облегчает проникновение в них вируса. Путем пассивного центростремительного продвижения по аксоплазме нервных волокон он достигает центральной нервной системы, где внедряется в нейроны дорсальных ганглиев спинного, а затем и головного мозга, накапливаясь в нейронах гиппокампа, ядер моста, мозжечка, лимбической системы и обонятельных луковиц. Из ЦНС вирус диссеминирует по всему организму тем же путем, но уже в центробежном направлении. Уже за несколько дней до появления первых клинических признаков он может выделяться в окружающую среду через респираторный тракт, мочевыделительную систему, со слюной. В связи с богатой иннервацией слюнных желез титр вириона в секрете ротовой полости намного превышает его содержание в мозговой ткани. Выход в протоки слюнных желез обеспечивает передачу вируса со слюной при укусах. В крови вирус не обнаружен.
Входными воротами являются не только клетки поперечнополосатых мышц, нервно-мышечные бляшки, проприорецепторы сухожилий, но и рецепторы вкусовых сосочков языка и нейроэпителий обонятельных луковиц. Известны, в частности, случаи бешенства у спелеологов, занимающихся исследованием пещер. Здесь обитает большое количество насекомоядных летучих мышей — бессимптомных носителей вируса бешенства. Со слюной мышей он выделяется в окружающую среду и при вдыхании зараженного аэрозоля может вызвать заболевание.
Скрытая персистенция уличного вируса бешенства (для нее характерен слабый цитопатический эффект и непрерывный, но очень низкий уровень продукции вируса) выявляется также у собак, песцов, волков, грызунов, летучих мышей. Их укусы могут приводить к заболеванию и смертельному исходу. В процессе персистенции возможны изменения различных свойств вируса, в том числе повышение вирулентности, что способствует формированию новых эпидемически значимых вирусных штаммов.
Различают следующие периоды заболевания:
- инкубационный,
- продромальный,
- развившейся болезни (стадия возбуждения),
- фазу параличей, неизбежно заканчивающуюся летальным исходом.
Инкубационный период бешенства чрезвычайно вариабелен: от 10 дней до года и более (в среднем 1—2 мес). Его продолжительность зависит от места внедрения вируса (наиболее короткий — при укусах в лицо, голову, шею, пальцы рук), заражающей дозы, степени тяжести и объема пораженной ткани, возраста пострадавшего (короче — у детей). Первые признаки болезни почти всегда обнаруживаются в месте укуса. Появляются тянущие, ноющие боли, зуд, невралгические боли по ходу нервных путей, ближайших к месту укуса. Затем развивается астения: общее недомогание, чувство дискомфорта, беспокойство, раздражительность. Явления беспокойства могут принимать агрессивный характер и даже перейти в состояние буйства. Присоединяются слуховые и зрительные галлюцинации, возникают пароксизмы (приступы) (гидро-, аэро-, фото- и акустикофобии), сопровождающиеся чрезвычайно болезненными спазмами мышц глотки. Отмечаются головная боль, субфебрильная температура, обильное слюноотделение. В результате обезвоживания черты лица заостряются, снижается масса тела. Если смерть не наступает во время очередного приступа, болезнь переходит в паралитический период. Ухудшается сердечно-сосудистая деятельность, появляются параличи мышц конечностей и черепно-мозговых нервов, дисфункции тазовых органов. Смерть наступает от паралича дыхательного и сосудистого центров.
Заболевание может протекать без явлений возбуждения в виде восходящего паралича — так называемое тихое бешенство. В таких случаях гидро- и аэрофобия отсутствуют или выражены слабо. Болезнь протекает дольше, смерть наступает вследствие бульбарных расстройств (паралич дыхательного и сосудистого центров).
У людей, погибших от бешенства, обнаруживаются воспалительные процессы в оболочке и тканях мозга, нервных ганглиях. Аналогичные изменения выявляются в слюнных железах и надпочечниках. В нейронах головного мозга возникают внутриклеточные, эозинофильные включения, состоящие из тонкого фибриллярного матрикса и вирусного рибонуклеокапсида. Они были выявлены еще в 1903 г. Негри при окраске среза продолговатого мозга метиленовой синькой и эозином («тельца Негри»). Считают, что это проявление одной из защитных реакций, ограничивающих распространение вируса в нервной ткани.
Другим лимитирующим механизмом может быть опережающий (до созревания вириона) апоптоз зараженных клеток. Оказалось, что не все вирусы возбуждают этот процесс, причем степень вирулентности обратно пропорциональна апоптозиндуцирующей способности. Высоковирулентные уличные (дикие) штаммы, циркулирующие в природе, вообще не стимулируют его.
Специфическая диагностика и профилактика
Классическим методом постмортальной диагностики служит выявление телец Негри в тканях мозга. Для этого используют специальное окрашивание или иммунофлюоресцентный метод. Кроме того, пассируя через мышей, вирус выделяют из слюнных желез, тканей мозга и затем заражают клеточные культуры. О присутствии вируса судят по минимальным цитопатическим изменениям или по определению вирусных антигенов в культуре тканей иммунофлюоресцентным методом.
Вирус можно выявить в слюне, мазках-отпечатках с конъюнктивы глаза, кожных биоптатах или определить его присутствие (цереброспинальная жидкость, слюна, слезная жидкость) при помощи полимеразной цепной реакции.
Методов терапии бешенства не существует. Прогноз — неблагоприятный, достоверных случаев выздоровления нет. Основное значение имеет опережающее применение антирабических иммунопрепаратов — вакцины и специфического иммуноглобулина. Лицам, получившим множественные укусы или повреждения опасной локализации, предварительно вводят антирабический иммуноглобулин, а затем проводят курс вакцинации. Антитела после прививок появляются лишь через 3 нед. При ранениях неопасной локализации проводят только вакцинацию. Иммунитет сохраняется на протяжении года.
Как уже говорилось, первая вакцина, предложенная Пастером, была получена путем пассирования вируса бешенства через мозг животных. «мозговая» вакцина использовалась длительное время, но иногда она дает тяжелые аутоиммунные поствакцинальные осложнения (энцефалиты и др.). В связи с этим в настоящее время используют культуральную вакцину — на основе вируса, адаптированного к культуре тканей.
Профилактика бешенства включает целый комплекс мероприятий. Обязательны регистрация и вакцинация домашних животных, примение жестких карантинных мер при их импорте, отлов и стерилизация бродячих собак. Имеется положительный опыт (у нас в стране и за рубежом) профилактики бешенства среди диких животных путем скармливания им приманок, содержащих вакцину. Перспективными выглядят рекомбинантные оральные вакцины с использованием в качестве вектора вируса осповакцины, экспрессирующего ген протективного G-протеина вируса бешенства.
Арбо- и родентвирусы
И комар лошадь свалит, коли волк пособит.
- Арбовирусы и арбовирусные инфекции:
тогавирусы: альфа-вирусы и вирус краснухи;
флавивирусы: количество и качество;
буньявирусы: вирусология против экологии.
- Родентвирусы и родентвирусные инфекции:
хантавирусы: ГЛПС и кардиопульмонарный синдром;
астравирусы: двойное доказательство природной очаговости.
Арбо- и родентвирусы представляют собой экологически близкие агенты, связанные с животными. Человек для них случайный хозяин, и его инфицирование гибельно для большинства из них. Однако арбовирусы устроены сложнее. Они нуждаются в дополнительном организме, которым являются кровососущие членистоногие, где происходит персистенция вирусов при длительном бездействии. Родентвирусы не нуждаются в таком резервуаре, передаваясь между животными и человеком без посредников. И все-таки мы сочли возможным говорить о родентвирусах рядом с арбовирусами для того, чтобы их различия не закрывали глаза на патогенетическую общность инфекций, вызываемых ими у человека.
Арбовирусы
Арбовирусы получили название от английских слов arthropod born viruses, что означает «вирусы, передающиеся членистоногими». Так называется группа инфекционных агентов, которые заражают человека и животных через укусы кровососущих членистоногих, главным образом комаров и клещей. Из позвоночных животных в жизни арбовирусов участвуют дикие птицы и грызуны, в меньшей степени — средние и крупные животные; определенную роль играют домашние животные (лошади, свиньи и др.).
В членистоногих арбовирусы проходят цикл размножения, заканчивающийся в слюнных железах. Это называется внешним периодом инкубации. Репликация в организме членистоногих проходит без признаков болезни. Более того, немало арбовирусов сохраняется в них, передаваясь трансовариальным и/или трансфазовым путем. Это позволяет вирусу выживать в условиях длительного бездействия, например в зимнее время. Членистоногие становятся не только переносчиком, но и резервуаром инфекции. Напротив, позвоночные могут болеть и умирать от арбовирусных инфекций, но чаще все для них складывается благополучно. Человек попадает в зоонозный цикл случайно, подвергаясь нападению зараженных членистоногих. Обычно он служит экологическим тупиком, из которого возбудитель не имеет эволюционного выхода.
Как правило, арбовирусы сконцентрированы в тропическом и субтропическом регионах. Это объясняется изобилием животных в этих зонах и благоприятными условиями для биологических циклов, которые включают кровососущих членистоногих и позвоночных. Но арбовирусы встречаются и в странах с умеренным климатом, в частности в России. Возможны привозные случаи инфекций, если учесть, что инкубационный период (он составляет в среднем 5—7 дней) допускает это при современных скоростях пассажирских перевозок.
Многим арбовирусным болезням присущи сезонные колебания. Они связаны с изменением природных факторов, которые отражаются главным образом на переносчиках инфекции. Первоочередным условием является температура, влияющая на размножение возбудителя. Например, при 37°С вирус желтой лихорадки в теле комара Aedes aegypti (основной переносчик) достигает инфицирующей дозы через 4 дня; при 18°С этого не происходит даже через 30 дней. Поэтому температурный режим служит важнейшим фактором, который ограничивает ареал желтой лихорадки: переносчик распространен значительно шире, чем сам вирус. Существенное влияние оказывают осадки и продолжительность сухого периода. Большое значение имеют трофические связи членистоногих с позвоночными. Многие комары питаются главным образом на птицах, тогда как другие виды используют для этого млекопитающих.
Названия некоторых арбовирусов отражают характер связанных с ними болезней — вирус желтой лихорадки, вирус денге и др. Позже вирусы стали называть по географической области, где они были выделены. Так появилось множество обозначений — вирусы энцефалита Сент-Луи, Омской лихорадки, Синдбис, Укуниеми, Чикунгунья, лихорадки Западного Нила, Конго-Крымской лихорадки, Карельской лихорадки, Калифорнийского энцефалита, Венесуэльского энцефалита и т.д. Сейчас известно около 500 арбовирусов, из них примерно 100 поражают человека.
Большинство арбовирусов принадежит семействам Togaviridae, Flaviviridae и Bunyaviridae. Единичные арбовирусы есть среди семейств Reoviridae, Rhabdoviridae и Picornaviridae. Все они являются РНК-вирусами. Индивидуальные серотипы (виды) Togaviridae, Flaviviridae и Bunyaviridae отличаются друг от друга по тесту нейтрализации с иммунными сыворотками. Для их определения пользуются моноклональными антителами, которые дифференцируют эпитопы, недоступные для поликлональных антител. Серотипы неоднородны. Это проявляется в реакциях торможения гемагглютинации, связывания комплемента и иммуноэнзимного анализа. Серотипы, дающие выраженные перекрестные реакции, объединены в антигенные комплексы, или серогруппы. На этом основании выделено 7 групп (комплексов) тогавирусов (альфавирусов), 8 — флавивирусов и 14 — буньявирусов. Антигенные связи отражают происхождение от единого источника, однако дальнейшее разделение происходит в 10—100 раз медленнее, чем у других РНК-вирусов, не связанных с членистоногими.
Выделяют три категории арбовирусных инфекций:
- лихорадки с развитием артралгий и сыпи;
- геморрагические лихорадки с кровоизлияниями в коже, слизистых оболочках и различных органах;
- энцефалитные синдромы.
Болезнь может протекать с гриппоподобными (неспецифическими) симптомами или вообще не развиваться, ограничиваясь образованием антивирусных антител. Таких случаев большинство, и в среднем свыше 90% заражений протекают бессимптомно. Для многих арбовирусов число инаппарантных форм составляет 300:1—1000:1, демонстрируя штаммовую неоднородность, зависимость от условий заражения и масштаб устойчивости к арбовирусам. К манифестным формам более чувствительны пожилые и дети. Один и тот же вирус может вызвать неодинаковые синдромы у разных людей.
Общая схема арбовирусных инфекций показана на рис. 1. Заболевания возникают после укуса членистоногих, слюна которых содержит инфицирующую дозу вируса. Проникая через кожу, вирусы реплицируются местно и в регионарных лимфоузлах, после чего поступают в кровь (первичная вирусемия). Высвобождение вируса из клеток селезенки, печени, лимфатических узлов и сосудистого эндотелия создает вторичную вирусемию и может спровоцировать гриппоподобную симптоматику. Через неделю (после появления достаточного количества нейтрализующих антител) вирусемия исчезает. Инфекция обычно на этом заканчивается, но иногда возбудитель успевает достичь дополнительных мишеней — центральной нервной системы, кожи, мышц, суставов и т.д. Способность разных клеток воспринимать и поддерживать вирусную инфекцию сильно отличается.

Между началом основного синдрома и неспецифической фазой может быть короткий промежуток облегчения, и тогда болезнь протекает в виде двух фаз. Специфические симптомы начинаются через 7—10 дней после заражения и (в зависимости от вируса и хозяина) продолжаются неделю или дольше, заканчиваясь выздоровлением, ремиссией или смертью. Геморрагический синдром сопровождается кровоизлияниями — мелкоточечной сыпью (петехии на коже и слизистых оболочках), кровотечением из носа, кровавой рвотой и калом, кровоизлияниями во внутренние органы. Скорее всего, это связано с тромбоцитопенией и диссеминированной коагулопатией. Смертность при геморрагических лихорадках колеблется от 1 до 15%, но может быть и выше.
При энцефалитах арбовирусы размножаются в нейронах головного мозга, вызывая образование зон некроза, окруженных микроглиальными клетками. Такие же скопления появляются вокруг многих церебральных сосудов. Одновременно возникает асептический менингит с аккумуляцией мононуклеаров в субарахноидальном пространстве и гиперемией прилежащих капилляров. Энцефалит (точнее менингоэнцефалит) начинается с подъема температуры, головной боли и ригидности шейных мышц. Прогрессируя, это ведет к спутанности сознания и неврологическим симптомам — тремору, патологическим рефлексам, парезам и параличам, нистагму, атаксии, конвульсиям, коме. Смертность достигает 80%. Нервные последствия (парезы и пр.) остаются у многих больных, но через несколько месяцев проходят. Нетрудоспособность, хроническая усталость, дефицит памяти и концентрации внимания могут сохраняться годами.
Большинство арбовирусов культивируются в различных клетках и патогенны для новорожденных мышей. Поэтому заражение культуры клеток и мышей остаются главными методами диагностики арбовирусных инфекций. Из других способов используется определение IgM- и IgG-антител и обратнотранскриптазная полимеразная цепная реакция — ПЦР (поиск вируса в циркуляции).
Для профилактики арбовирусных инфекций большое значение имеют экологические подходы, прежде всего борьба с комарами и клещами. При некоторых инфекциях применяют убитые или живые аттенуированные вакцины.
Семейство тогавирусов
Семейство Togaviridae (от лат. toga — плащ) включает два экологически обособленных рода — альфавирусы (Alphavirus) и рубивирус (Rubivirus). Альфавирусы — типичные арбовирусы, передающиеся через кровососущих членистоногих. Рубивирус, или вирус краснухи, представляет собой сугубо человеческий вирус; инфицирование происходит воздушно-капельным путем, не требуя участия кровососущих переносчиков. Ниже речь пойдет об альфавирусах, ибо вирус краснухи не относится к арбовирусам.
Вирионы являются сферическими образованиями, 60—70 нм в диаметре, с кубическим (икосаэдрическим) нуклеокапсидом, заключенным в липидно-белковую оболочку (суперкапсид). Геномной молекулой служит однонитчатая РНК длиной около 12 кбайт. Она действует как мРНК и обладает способностью самостоятельно инфицировать клетки, т.е. является плюс-РНК. Капсид образован С-белком (от англ. capsid); в суперкапсиде имеются короткие шипы, состоящие из гликопротеинов Е1 и Е2 (от англ. envelope). Третий протеин (Е3) удается обнаружить только у вируса леса Семлики; в остальных случаях он теряется в виде растворимого белка. Е1 определяет способность альфавирусов агглютинировать эритроциты птиц (гусей) и перекрестно реагировать в реакции торможения гемагглютинации (для штаммов одного антигенного комплекса). Е2 тоже вызывает гемагглютинацию, но она более специфична, чем у Е1. К тому же Е2 служит мишенью для антител, нейтрализующих альфавирусы. Нейтрализующие тесты определяют разделение вирусов на типы или виды внутри антигенных комплексов.
С мутациями в генах, кодирующих Е1 и Е2, связаны колебания вирулентности тогавирусов (альфавирусов), что в известной мере объясняется изменением рецепторной активности вирионов. Идентифицировано несколько клеточных белков, реагирующих с альфавирусами. Степень их аффинности и функциональной активности различна, но, действуя вместе, они обеспечивают поглощение вириона. Дальнейшие события происходят в клеточной цитоплазме. В кислой среде вирион, используя сайты слияния Е1, образует стыковку между суперкапсидом и эндосомальной мембраной. Это ведет к высвобождению РНК, которая, действуя как мРНК, производит полипротеин для четырех неструктурных белков, в том числе для РНК-зависимой РНК-полимеразы. РНК-полимераза обеспечивает продукцию новых копий вирионной и субгеномной РНК, дающей начало структурным (вирионным) белкам. Они транспортируются в плазматическую мембрану; сюда же подходят готовые нуклеокапсиды. После связывания Е2- и С-белков происходит почкование новых вирионов и разрушение клетки. В культуре тканей полная деструкция клеточных монослоев наступает через 48—72 ч.
Известно около 30 видов альфавирусов, половина из которых опасна для человека. По особенностям своих поверхностных гликопротеинов альфавирусы разделены на семь антигенных комплексов (групп), главными среди которых являются комплексы Восточного лошадиного энцефалита, Венесуэльского лошадиного энцефалита, Синдбис (включает вирус Западного лошадиного энцефалита) и леса Семлики. Все альфавирусы передаются комарами и в большей степени связаны с дикими птицами. Некоторые ассоциированы с птицами и грызунами, другие — только с грызунами или с приматами. Человек является экологическим тупиком почти для всех альфавирусов; исключение составляют представители комплекса леса Семлики — вирусы Чикунгунья, ОўНъонг-Нъонг (Африка, Азия) и Росс Ривер (Австралия). Эти вирусы приспособились к синантропным (домашним) комарам (разновидности Aedes, Anopheles, Culex), которые нападают на больных людей в фазу вирусемии, запуская цикл «человек—комар—человек», что ведет к эпидемическим вспышкам в густонаселенных районах.
Главные альфавирусные заболевания человека — тяжелые лихорадки (нередко осложненные сыпью и артритами) и энцефалиты. В инкубационном периоде и ранней клинической стадии они сопровождаются присутствием вируса в крови (вирусемией). Многие случаи ограничиваются неспецифическими (гриппоподобными) симптомами или вообще протекают без клинических признаков на фоне образования антител.
Большинство инфекций регистрируется в Центральной и Южной Америке (комплексы Западного, Восточного и Венесуэльского лошадиных энцефалитов, вирусы группы Синдбис и леса Семлики). Меньше страдает население Юго-Восточной Азии, Африки и Австралии. Лихорадка Синдбис зарегистрирована в южных районах Европы.
Для профилактики кроме борьбы с комарами используются убитые формалином вакцины, которые дают высокую степень защиты (вирусы Западного, Восточного и Венесуэльского лошадиных энцефалитов). Вакцины применяют по эпидемиологическим показаниям — лабораторные служащие, лица с высоким риском инфицирования.
Анализ инфекций строится на основании общих принципов, принятых для диагностики арбовирусных заболеваний: выделение из крови вируса-возбудителя, определение IgM- и IgG-антител, поиск вирусных РНК методом обратнотранскриптазной ПЦР.
Семейство флавивирусов
Cемейство Flaviviridae получило название от желтой (лат. flavus) лихорадки, которая в прошлом относилась к числу наиболее опасных болезней Центральной и Южной Америки. Это первая инфекция человека, для которой В. Ридом в 1901 г. была установлена вирусная природа. В дальнейшем обнаружено множество родственных вирусов, которые по началу были включены в семейство Togaviridae и обозначены как группа В, чтобы отличить их от группы А, теперь известной как альфавирусы (см. выше). В 1984 г. введен самостоятельный таксон — семейство Flaviviridae. В нем учитываются новые данные о генетических и фенотипических свойствах этих вирусов.
Флавивирусы — наиболее важная в медицинском отношении группа арбовирусов. К ним относится возбудитель лихорадки денге — самой распространенной инфекции человека тропического пояса. Эпидемической угрозой для Африки и Южной Америки остается желтая лихорадка. Энцефалиты, переносимые комарами и клещами, представляют опасность для Америки, Европы и Азии. Сложные циклы трансмиссии и экологической перестройки наряду с пластичностью РНК-генома ставят флавивирусные болезни в число «вновь появляющихся инфекций» (англ. emerging and reemerging infections). Примером служат Омская геморрагическая лихорадка, энцефалит Западного Нила, желтая лихорадка и др.
Флавивирусы включают вирусы трех родов — Flavivirus, Pestivirus и Hepacivirus. К арбовирусам человека принадлежат только представители рода Flavivirus (собственно флавивирусы). К пестивирусам относятся возбудители ветеринарных инфекций; род Hepacivirus состоит из вирусов гепатита С и G человека (см. «Вирусы гепатитов»).
Род Flavivirus cодержит более 70 вирусов. Большинство из них связаны с комарами и клещами; более половины поражают человека. По результатам серологической нейтрализации, подтвержденной генетическим анализом, флавивирусы разделены на виды (типы), которые на основании реакции торможения гемагглютинации объединены в группы (комплексы). Новые возможности открывает использование моноклональных антител. С их помощью удается обнаружить родовые, групповые и специфические эпитопы оболочечного гликопротеина Е. Известно по меньшей мере восемь антигенных комплексов. Наибольшее число вирусов представлено комплексом клещевого и японского энцефалитов.
Вирион представляет собой частицы размером 40—50 нм. Внутри расположен нуклеокапсид кубической симметрии, снаружи — липидная оболочка (суперкапсид), в которой имеются небольшие выступы, состоящие из Е-гликопротеина. Это единственный компонент, отчетливо выявляемый на поверхности вириона. Он определяет вирусную способность к гемагглютинации, рецепции, слиянию с клеточными мембранами и взаимодействию с нейтрализующими антителами. Кроме Е-гликопротеина, вирион располагает внутренним С-белком, который образует капсид, связанный с геномной молекулой. В липидной оболочке присутствует еще один белок (М-протеин); в зрелом вирионе он почти незаметен. М-протеин выполняет цементирующую роль, стыкуя капсид с суперкапсидом. В процессе онтогенеза образуется пре-М-белок (prеM), который помогает созреванию и экзоцитозу вируса. Перед высвобождением вириона из клетки prеM подвергается протеолизу с удалением основного фрагмента. Это оставляет в суперкапсиде стабилизирующий М-белок.
Геном представлен линейной однонитчатой плюс-РНК размером около 11 кбайт. Вирионная РНК служит мРНК для всех вирусных белков и обладает инфекционностью при введении в чувствительные клетки. Гены структурных белков (С, preM и Е) cосредоточены на одном (5’) конце, за ними находятся гены для 7 неструктурных белков (NS1, NS2a, NS2b, NS3, NS4a, NS4b, NS5). Все белки синтезируются в виде полипротеина, который нарезается на индивидуальные компоненты NS3-cерин-протеазой. NS5-белок выполняет функции РНК-зависимой РНК-полимеразы, используя вирионную РНК как цельную матрицу. Функции остальных неструктурных белков не известны.
Репликация флавивирусов начинается с Е-зависимого прикрепления к гликозаминогликанам и, возможно, другим (пока не идентифицированным) рецепторам плазматической мембраны. Так же, как в случае альфавирусов, это ведет к эндоцитозу вирионов. Нуклеокапсиды высвобождаются в результате слияния Е-белка с эндосомальной мембраной, давая начало трансляции геномной РНК. Одновременно образуется РНК-матрица для синтеза новых вирионных РНК и продукции структурных вирусных белков. Зрелые вирионы обнаруживаются внутри цитоплазматических вакуолей и высвобождаются при их слиянии с плазматической мембраной клетки; почкования не происходит. Размножение флавивирусов протекает медленнее, чем альфавирусов, их действие не носит выраженного цитопатического эффекта, а синтез белков клетки-хозяина происходит вместе с продукцией вируса.
Механизмы патологии широко варьируют. Безопасные для переносчиков, флавивирусы могут вызывать болезни у позвоночных животных, в том числе у человека. Говоря о болезнетворности для людей, следует выделить вирусы энцефалита, лихорадки денге и желтой лихорадки.
Энцефалитные вирусы переносятся комарами Culex и Aedes (энцефалит Сент Луи — Северная, Центральная и Южная Америка; Японский энцефалит — Индия, Китай, Япония, Юго-Восточная Азия; энцефалит долины Маррей — Австралия) и иксодовыми клещами Ixodes persulcatus и Ixodes ricinus (Европа, Азия). Вирусы клещевого энцефалита способны перезимовывать в теле клеща и передаваться трансовариально и трансфазово (яйцо—личинка—нимфа—имаго). В России заболевания регистрируются на всей лесной территории — от Дальнего Востока до западных границ. Прокормителями клещей и вторым по значимости резервуаром инфекции служат мышевидные грызуны, ежи, белки, бурундуки, кроты, крупные млекопитающие, мелкий и крупный рогатый скот. Различают западный (европейский) и восточный (сибирский) клещевые энцефалиты, которые вызываются двумя антигенными субтипами одного и того же вируса, передаются разными клещами (I. ricinus — западный и I. persulcatus — восточный вариант) и отличаются по нейровирулентности (восточный энцефалит протекает тяжелее). Заражение возможно при употреблении сырого молока инфицированных коз и коров. Кстати, козы болеют клещевым энцефалитом; коровы устойчивы к вирусу, но у них тоже наблюдается вирусемия. Вирус может длительно персистировать в центральной нервной системе, давая клинические проявления прогрессирующего характера. Для профилактики предложена убитая формалином вакцина.
Из энцефалитных флавивирусов остановимся еще на одном вирусе — вирусе лихорадки Западного Нила (ЛЗН). Он был изолирован в 1937 г. в провинции Западный Нил (Уганда) от больного с легкой формой лихорадки. В дальнейшем оказалось, что тот же вирус способен вызывать тяжелые поражения центральной нервной системы не только в Африке, но и в Южной Европе, на Среднем Востоке, в Южной Азии и Австралии. В 1999 г. неожиданно возникла вспышка в США (г. Нью-Йорк и его окрестности), которая сопровождалась высокой гибелью мигрирующих птиц. В России примерно в это же время зарегистрирована вспышка ЛЗН в южных регионах — Астраханской, Волгоградской областях и Краснодарском крае. Дело в том, что основной цикл вируса проходит между различными птицами и орнитофильными комарами рода Culex, среди которых есть виды, охотно нападающие на человека. Больше всего комаров наблюдается в июле—августе, и именно в это время происходит подъем заболеваемости среди людей. Человеческим вспышкам предшествуют эпизоотии среди диких, а затем и домашних птиц. С птицами связан и межконтинентальный обмен вирусными популяциями ЛЗН.
Заражение ЛЗН часто не ведет к клиническим симптомам, хотя и сопровождается образованием антител. В клинически значимых случаях (чаще пожилые люди и дети) после 3—6-дневной инкубации возникают гриппоподобные симптомы и развивается генерализованная лимфаденопатия. У половины больных на туловище и верхних конечностях появляется розеолезная или макулопапулезная сыпь. Заболевание обычно заканчивается через 3—6 дней. Иногда наблюдается поражение ЦНС (асептический менингит и менингоэнцефалит); еще реже выявляются экстраневральные осложнения (гепатит, миокардит, панкреатит). В таких случаях смертность достигает 10%. Вакцины нет, и приходится полагаться на неспецифические меры профилактики.
Лихорадка денге — самая распространенная арбовирусная инфекция человека в тропическом и субтропическом климате. Встречается более чем в 60 странах с населением свыше 2,5 млрд. человек. Ежегодно наблюдается более 100 млн. случаев болезни, 100—200 человек возвращаются больными в США, Европу и Австралию из поездок в эндемичные районы.
В городских условиях заболевание передается через укус синантропного комара, Aedes aegypt — возникает антропонозная форма заболевания. Комары заражаются от человека во время вирусемии (инкубационный период и первые дни болезни) и становятся опасными через 8—14 дней после кровососания. Эпидемии наступают после сезона дождей, когда увеличивается численность комара-переносчика. Следующему поколению комаров вирус не передается.
Кроме антропоноза, существует зоонозная форма — лихорадка джунглей. Резервуаром вируса служат обезьяны; переносчиками — комары рода Aedes (A. albopicans и A. niveus), переносящие возбудителя от животных человеку.
Заболевание вызывают четыре антигенных типа вируса — 1, 2, 3 и 4. Зависимость между вирусным типом и клинической картиной отсутствует. Вирус начинает размножение в дендритных клетках или макрофагах, которые накапливаются в месте укуса. Отсюда он проникает в регионарные лимфатические узлы и отдаленные ретикулоэндотелиальные органы. Вирусемия (как и при всех арбовирусных инфекциях) — обязательный элемент патогенеза. Она заканчивается с появлением вируснейтрализующих антител.
В 1869 г. Королевский Колледж врачей в Лондоне ввел единый термин — лихорадка денге. «Денге» заимствовано из фразы на языке суахили «ka denga pepo», что означает «внезапное появление болей у пациента». Речь идет о тяжелейшем болевом синдроме в мышцах, суставах и спине. Отсюда старое, но до сих распространенное название — костоломная лихорадка (англ. breakbone fever).
Через 3—7 дней после заражения среди полного здоровья появляются озноб, быстрое повышение температуры (39—40°С), нарастающие боли в спине, крестце, суставах. Наблюдаются тошнота, рвота, анорексия, головокружение, бессонница, гиперемия и пастозность лица, инъекция сосудов склер, гиперемия зева. Лихорадка обычно падает к концу третьих суток, но температура вновь повышается через 1—3 дня. Возникает полиморфная сыпь, чаще макулопапулезного (кореподобного) характера. Повторной волне заболевания сопутствует генерализованная лимфаденопатия, гранулоцитопения и тромбоцитопения. Все это характерно для классической формы денге. Она наблюдается у 1 из 6—15 человек, подвергшихся первичному заражению и, как правило, заканчивается самоизлечением через 3—9 дней.
Иначе протекает повторный контакт с гетерологичным типом вируса, особенно со вторым типом. Это чаще наблюдается в детском возрасте через 3 мес — 5 лет после первичной инфекции. Возникает геморрагическая лихорадка, которую называют денге-шоковым синдромом. Поражение эндотелия мелких сосудов ведет к геморрагическому диатезу в коже, слизистых оболочках и внутренних органах, аноксии тканей и метаболическому ацидозу. Утечка плазмы из капилляров сопровождается гиповолемическим шоком — похолоданию кожи, цианозу, учащению пульса, сонливости. В 25—50% случаев наступает смерть. Выжившие быстро поправляются.
Есть мнение, что патогенез повторной лихорадки связан с иммунопатологическими процессами, но пока трудно судить о правомерности предполагаемых механизмов, тем более что сходные симптомы возникают не при каждой вторичной инфекции. Другая версия допускает генетическую изменчивость вируса, которая увеличивает его патогенетический потенциал, в частности репликацию и вирусемию.
До сих пор не существует вакцины против денге. Многие способы приготовления вакцинных препаратов проходят проверку, начиная от цельновирионных до «голых» ДНК-вакцин. Отметим, например, многообещающую попытку создания препарата на основе вакцинных клонов вируса желтой лихорадки или Японского энцефалита с включением в них генов для структурных белков вирусов денге.
Вирус желтой лихорадки вызывает геморрагический синдром с поражением печени, почек и миокарда. Передается комарами и встречается в тропическом и субтропическом поясах Южной и Центральной Америки, Западной, Центральной и Восточной Африки. Ранее это была одна из самых страшных эпидемических болезней и неоднократно заносилась в южные страны Европы и Северную Америку. В годы строительства Панамского канала от желтой лихорадки погибло около 10000 человек. На протяжении последних 40 лет вспышки заболевания с высокой летальностью возникали в Судане, Эфиопии, Сенегале, Нигерии, Анголе и других странах Африки и Южной Америки. Коэффициент инфекция/болезнь колебался от 2:1 до 20:1.
В разных регионах встречаются по крайней мере пять Е-генотипов вируса желтой лихорадки. Гомология между ними превышает 90%, доказывая слабую мутационную изменчивость вируса, который, как полагают, возник раньше других флавивирусов — около 3000 лет назад. Подобно лихорадке денге, имеется две взаимосвязанных формы заболевания — лихорадка джунглей и лихорадка городского типа. Первая — типичный зооноз с природной очаговостью, лихорадка городского типа — антропоноз, переносимый от человека к человеку синантропными комарами. Антропонозные очаги создаются людьми, работающими в джунглях. Резервуаром служат обезьяны. Они безболезненно переносят контакт с вирусом, но создают вирусемию, достаточную для заражения комаров. В Африке такие очаги поддерживаются комарами рода Aedes, в том числе A. luteocephalus, A. furcifer, A. taylori, нападающими на людей. В Америке основным переносчиком являются комары рода Haemagogus. Они атакуют людей не только в джунглях, но и в жилых постройках.
При лихорадке городского типа переносчиком является синантропный комар A. aegypti. Комар сохраняет вирус пожизненно, но трансовариально его не передает. При температуре ниже 20оС вирус не воспроизводится, однако при повышении температуры вновь начинает размножаться, проникает в слюнные железы, и через 4—12 дней комар становится инфекционно опасным.
Патогенез типичен для геморрагических лихорадок. Вирус первоначально размножается в регионарных лимфоузлах, а затем через кровь попадает в другие органы, где продолжает репликацию. При легких формах наблюдаются повышение температуры, головная боль и миалгия. Желтуха и альбуминурия отсутствуют. Типичная желтая лихорадка имеет двухфазное течение. Первая фаза начинается после короткого (3—6 дней) инкубационного периода, совпадая с вирусемией. Быстро поднимается температура, возникает головокружение, тошнота, боль в спине. Вслед за ремиссией наступает вторая фаза и проявляются главные симптомы болезни. Температура вновь повышается, поражение печени ведет к желтухе (отсюда желтая лихорадка), поражение почек — к выраженной альбуминурии и острой почечной недостаточности, появляется геморрагическая сыпь, кровавая рвота, кровотечения из носа, матки, мочевого пузыря, кишечника. Смерть наступает у 20—50% больных на 7—10-й день болезни. Бывают и молниеносные формы. Они с первого дня болезни сопровождаются кровавой рвотой; больные погибают до возникновения желтухи от острой сердечной недостаточности, желудочно-кишечных кровотечений, шока.
Для специфической профилактики используется вакцина из живого авирулентного штамма 17D, который получен в 1936 г. путем многократных пассажей вируса через организм мышей, тканевые культуры и куриные эмбрионы. Протективный иммунитет (об этом судят по нейтрализующим антителам) возникает у 90—99% вакцинированных и сохраняется по крайней мере на протяжении 20 лет.
Семейство буньявирусов
Буньявирусы — самое крупное семейство вирусов животных, включая человека. Обычно патология сводится к лихорадкам, не представляющим опасности, но иногда возникают тяжелые болезни, такие как Калифорнийский энцефалит, Конго-Крымская геморрагическая лихорадка, лихорадка долины Рифт и, наконец, хантавирусные геморрагические лихорадки с почечным и кардиопульмональным синдромами. Хантавирусные инфекции относятся к родентвирусным заболеваниям, и о них речь пойдет ниже.
Буньявирусы, вызывающие арбовирусные инфекции, включают три рода: Bunyavirus, Phlebovirus и Nairovirus. Они объединяют более 300 вирусов, разделенных на 14 серогрупп (комплексов), которые включают более 50 агентов, опасных для человека. Буньявирусы связаны с членистоногими переносчиками, причем трансовариальная и половая передачи превращают многих из них в резервуар инфекции. Большинство представителей рода Bunyavirus передаются комарами. Флебовирусы ассоциированы с москитами, найровирусы адаптировались к иксодовым и арагосовым клещам. Круг позвоночных хозяев весьма широк. Многие буньявирусы (речь идет о семействе) связаны с грызунами и зайцеобразными, меньше — с птицами, жвачными и другими домашними животными. Ряд вирусов ассоциирован с сумчатыми, летучими мышами, приматами и др. Привязка к определенной местности зависит от ограниченного распространения экологической системы, где существует вирус.
Буньявирусы являются оболочечными вирусами обычно круглой формы, размером 80—120 нм с гликопротеиновыми шипами длиной 5—10 нм. Белковолипидная оболочка (суперкапсид) окружает сердцевину, состоящую из трех спиральных нуклеокапсидов — короткого (S, от англ. short), среднего (M, middle) и длинного (L, long). К ним примыкают молекулы РНК-зависимой РНК-полимеразы, которая инициирует инфекцию. Это необходимо, так как геном буньявирусов представлен РНК (односпиральной) с негативной полярностью. Длина РНК-сегментов варьирует от 1 (S-сегмент) до 12 (L-сегмент) кбайт.
Единственным L-протеином является РНК-зависимая РНК-полимераза. М-сегмент кодирует поверхностные гликопротеины G1 и G2 (от англ. glycoprotein), а также неструктурный белок NSm. S-сегмент отвечает за синтез нуклеокапсидного белка N и неструктурного белка NSs.
Разные буньявирусы отличаются особенностями своего репликационного цикла. Вписываясь в общую картину размножения семейства, они придают своеобразие этой и без того разнородной группе вирусов. Рецепция и последующее взаимодействие с клеткой совершаются при участии одного или двух суперкапсидных гликопротеинов. После конформационных изменений G1 в кислой среде эндосом происходит слияние вирионной оболочки с эндосомальной мембраной и выход нуклеокапсида в цитоплазму. Минус-РНК используется для синтеза мРНК и образования вирусных белков. Одновременно это дает начало новым геномным минус-РНК. Возможна генетическая перетасовка между буньявирусами, принадлежащими к одной, но не к разным серогруппам.
Все события ограничиваются цитоплазмой. После сборки вирионных частиц наблюдается транспорт и слияние вируссодержащих везикул с плазматической мембраной и, наконец, выделение вирионов путем экзоцитоза. Схема напоминает флавивирусы за исключением того, что здесь отсутствует матриксный (М) протеин, сопрягающий суперкапсидные и нуклеокапсидные белки во время сборки вириона. Эту роль выполняет прямое взаимодействие оболочечных гликопротеинов с вирусными нуклеокапсидами.
Наибольшее значение имеют буньявирусы комплекса Калифорния (род Bunyavirus), которые включают более 10 серотипов (видов).
Из них выделим два наиболее распространенных вируса — Ла Кросс и Тягиня. Первый экологически связан с мелкими грызунами (бурундуки, белки), зайцами, кроликами Северной Америки. Переносчиками являются комары с трансовариальной и половой передачей вируса. Основным переносчиком вируса Ла Кросс для человека служит Aides triseriatus, который плодится в дуплах деревьев. Инкубационный период составляет 3—6 дней. Клиническая картина варьирует от лихорадочного синдрома до энцефалита. Болеют чаще дети 5—10 лет. Они лишены иммунной прослойки, которая появляется с ежегодным приростом около 1%. В эндемичных зонах к 60 годам примерно 20% населения обретает серопозитивность. Число смертельных исходов — менее 1%.
Вирус Тягиня, впервые выделенный в Африке, широко распространен в странах Западной, Центральной, Южной и Восточной Европы. Он изолирован также в северных областях Сибири, Дальнего Востока и Европейской части России, включая Заполярье. В Северной Европе циркулирует близкий вирус Инко. Среди позвоночных животных в поддержании природных очагов участвуют зайцы, ежи, лисы, белки и др., а из домашних животных — главным образом кролики и свиньи. Допускается подключение птиц, чем можно объяснить огромный ареал вируса в Европе, Азии и Африке. Вирус изолирован от 13 видов комаров, преимущественно рода Aedes, но для человека основным переносчиком является A. vexans, зараженность которого в Средней Европе составляет 1:1700. Лихорадка Тягиня обычно протекает как гриппоподобное заболевание. Многие случаи заражения фиксируются только по иммунным сдвигам.
Важными заболеваниями являются лихорадки, вызываемые сицилийским, неаполитанским вирусами и вирусом долины Рифт. Все они принадлежат к роду Phlebovirus и относятся к комплексу флеботомных лихорадок, так как кроме комаров передаются москитами (Phlebotomidae). Флеботомная лихорадка, связанная с сицилийским и неаполитанским вирусами, называется москитной лихорадкой, или лихорадкой паппатачи (итал. pape — обжираться и taci — молча). В передаче вируса участвуют многие виды москитов, среди которых главным является Phlebotomas papatasi. При температуре выше 18оС москиты обретают инфекционность спустя 7—10 дней и служат не только переносчиками, но и резервуаром вируса. Накапливаясь в яичниках москитных самок, возбудитель проникает в яйцеклетки, обеспечивая трансовариальную передачу вируса. Особенно важны антропонозный (человек—Ph.papatasi—человек) и синантропный циклы (домовая мышь—Ph. papatasi—человек—Ph. рapatasi—человек), которые могут привести к крупным вспышкам в густо населенных районах. Природные очаги (они связаны главным образом с песчанками) представляют опасность для людей, вступающих в контакт с зараженными москитами (геологи, охотники и др.). Заболевание наступает внезапно, через 3—9 дней после укуса, и протекает как лихорадка, иногда с менингеальным синдромом и повторными приступами. Прогноз обычно благоприятный. Патогенез и профилактика повторяют то, что известно для других арбовирусных инфекций. Вакцинация не разработана.
Лихорадка долины Рифт впервые описана в 1950—1951 гг. во время вспышки в долине реки Рифт в Кении (Восточная Африка). Тогда погибло более 100000 голов скота и заболело около 20000 человек. Природные очаги находятся в странах Южной, Западной и Восточной Африки. Однако в 1977 и 1993 гг. две вспышки произошли в Египте; вирус был занесен сюда верблюдами из энзоотичных территорий. Не исключена возможность распространения заболевания и на другие регионы, особенно на страны Средиземноморского бассейна и Юго-Западной Азии. Обычно болезнь носит доброкачественный характер с повышением температуры, миалгией, слабостью. Инкубационный период составляет 2—6 дней, субклинические случаи нетипичны. Симптомы проходят через 2—5 дней. Вместе с тем у 1% больных возникает геморрагический синдром с менингоэнцефалитными проявлениями, иногда с полной потерей зрения и гепатитом. Летальность — 0,25—0,5%, при тяжелых формах — 50%. Резервуаром возбудителя служат дикие и домашние животные (антилопы, верблюды, овцы, крупный и мелкий рогатый скот), а также комары (Culex, Aedes и Mansonia) и москиты, передающие вирус трансовариально. Для специфической профилактики применяется живая (для животных) и убитая формалином (для человека) вакцина.
Вирус Конго-Крымской лихорадки (ККЛ) относится к роду Nairovirus и принадлежит к числу наиболее значимых для России и стран бывшего СССР. Ареал ККЛ охватывает южные районы России, Азербайджан, Армению, Среднеазиатские страны, Молдавию, Крым. Циркуляция вируса установлена также в Европе (Болгария, Румыния, Греция, балканские страны и др.), Африке (Нигерия, Заир, Уганда) и Азии (Иран, Пакистан, Ирак, КНР и др.). История открытия вируса связана с именами российских ученых (М.П. Чумаков и соавт., 1945), которые сначала доказали вирусную природу заболевания (эпидемия геморрагической лихорадки наблюдалась после освобождения Крыма от фашистских захватчиков), а затем изолировали вирус от больного человека. В 1956 г. идентичный или весьма сходный вирус был выделен в Африке и получил название вируса Конго.
Вспышка ККЛ в Крыму была связана с возросшей популяцией зайцев и иксодовых клещей рода Hyalomma. Оказалось, что клещи Hyalomma являются не только переносчиком, но и хозяином вируса, поддерживая его трансфазово и трансовариально. Дополнительными хозяевами служат домашние животные (мелкий и крупный рогатый скот), зайцы и другие травоядные. Птицы, являясь прокормителями клещей Hyalomma, тоже участвуют в распространении вируса. Заболевание имеет выраженную сезонность, чаще возникая в весенне-летний период, т.е. во время активного нападения клещей Hyalomma. В отличие от большинства арбовирусов стертые и инаппарантные инфекции наблюдаются редко. Поэтому иммунная прослойка в природных очагах не превышает числа зарегистрированных больных. Инкубационный период составляет 2—12 дней. Вирусемия достигает пика в первые три дня болезни. Примерно в половине случаев заболевание имеет двухволновое течение: вслед за кратковременным снижением температуры наступает новый подъем и затем, начиная с 6—7-го дня, быстрое снижение и выздоровление. Этому предшествует геморрагический синдром, который проявляется в виде петехиальной сыпи, а у 10—15% больных более серьезными симптомами — носовыми кровотечениями, кровавой рвотой, гематурией, кишечными кровотечениями и пр. Летальность составляет 8—12%. Разработана инактивированная вакцина, но она скорее всего ненадежна, так как дает низкие титры вируснейтрализующих антител.
Родентвирусы
К родентвирусам относятся, как минимум, две группы вирусов — хантавирусы (род Наntavirus, семейство Bunyaviridae) и аренавирусы (представители семейства Arenaviridae). Их главным отличием от арбовирусов является то, что они лишены вектора членистоногих. Бессимптомная персистенция родентвирусов происходит внутри мышевидных грызунов (англ. rodents — грызуны). Вирусы выделяются с испражнениями и слюной, которые, загрязняя внешнюю среду, служат источником заражения новых особей, включая человека. Сохранение родентвирусов в природе поддерживается путем внутривидового/межвидового контакта позвоночных хозяев без участия членистоногих переносчиков.
Род хантавирусов (семейство буньявирусов)
Вирусология хантавирусов с некоторыми особенностями повторяет то, что известно для семейства буньявирусов. Например, у них отсутствуют неструктурные белки NSm и NSs, что, по-видимому, важно для взаимоотношений с клетками позвоночных. Геморрагическая лихорадка с почечным синдромом (ГЛПС) — первое и главное заболевание, связанное с хантавирусами. Больные с таким диагнозом встречаются на территории Европы и Азии. Много случаев наблюдается в Китае, Корее и на Дальнем Востоке. Ежегодно госпитализируется более 100000 больных, причем 40000—100000 из них только в одном Китае.
Первый штамм хантавирусов (вирус Хантан) был выделен в 1976—1978 гг. от полевых мышей Apodemus agrarius в эндемичном по ГЛПС районе недалеко от реки Хантан в Южной Корее. Позже было изолировано еще несколько антигенных вариантов вируса — Сеул, Добрава, Пуаамала и др. Они связаны с разными грызунами, имеют неодинаковое географическое распространение, сезонность и вызывают различные по тяжести заболевания.
Самые тяжелые формы ГЛПС вызывает вирус Хантан. Они встречаются в сельских районах Кореи, Китая и восточной части России. В типичных случаях после инкубационного периода 1—2 нед (реже до 4 нед) поднимается температура и возникает гриппоподобный синдром. Через несколько дней появляются тошнота, рвота и абдоминальные боли с генерализованной петехиальной сыпью и внутренними кровоизлияниями. Треть смертельных исходов происходит от шока в этом периоде. Затем наступает олигоуретическая стадия. Она продолжается в среднем 8 дней, сопровождается множественными кровоизлияниями в почечных капиллярах и ведет к острой почечной недостаточности. Примерно треть смертельных случаев наблюдается в этой фазе. Болезнь завершается диуретической стадией, когда больной ежедневно выделяет 3—6 л мочи. При легких формах этот период быстро проходит, но для трети больных может закончиться смертельным исходом. Общая летальность составляет 5—7%. Отношение инаппарантных к клинически выраженным формам составляет 14:1.
Другие хантавирусы вызывают умеренные формы ГЛПС. Это прежде всего касается вируса Пуаамала (хозяин — красная полевка), ответственного за эпидемическую нефропатию. Она наблюдается в Скандинавии, странах Западной и Восточной Европы, в Европейской части России. Смертность составляет около 0,2%.
Cледует отдельно сказать о вирусе Сеул. В отличие от большинства ГЛПС, которые наблюдаются в сельских местностях при контакте с выделениями диких грызунов, этот вирус связан с синантропными крысами и преимущественно распространен в городах. Вирус Сеул встречается во всем мире, но взаимодействие с ним чаще не имеет последствий, кроме образования антител.
Полагают, что хантавирусы обладают тропизмом к почечной ткани, хотя прямые доказательства этого отсутствуют. Не исключено, что речь идет об иммунологически опосредованном нарушении эндотелиальных клеток, которое определяет утечку плазмы и эритроцитов. Инфекция не передается между людьми и ведет к длительному (возможно, пожизненному) иммунитету. Имеются сообщения о двух формалинизированных вакцинах (вирусы Хантан и Сеул), лицензированных в Корее и Китае.
Второе заболевание описано в 1993 г. среди сельских жителей США. Это тяжелый, часто смертельный кардиопульмонарный синдром. Возбудителем оказался новый серотип хантавирусов, получивший название «Sin Nombre вирус кардиопульмонарного синдрома» (SNV). Его главным хозяином служит оленья мышь (Peromyscus maniculatus), широко распространенная в Северной Америке. Человек заражается, вдыхая пыль с экскрементами инфицированных грызунов. Вирус обладает тропизмом к легочной ткани, вызывая тяжелый респираторный дистресс-синдром с типичным отеком легких, возможно, из-за повышения проницаемости микрососудов. Кровоизлияния отсутствуют, хотя, подобно ГЛПС, синдром сопровождается тромбоцитопенией. Случаи пульмонарного синдрома наблюдали в Центральной и Южной Америке, причем численность вирусных серотипов возросла до 11. К счастью, ни один из них не вышел за границы Америки. Инфекция не передается от человека к человеку.
Семейство аренавирусов
Название семейства происходит от лат. arena — песок. Это связано с тем, что во время репликации вирусные частицы обретают электроноплотные зерна клеточных рибосом, которые напоминают гранулы песка. Из 15 известных аренавирусов только пять вызывают болезни у человека. Три из них встречаются в странах Южной Америки (вирусы Хунин, Мачупо, Гуйнарио) и называются вирусами Нового Света. Два — получили название вирусов Старого Света; они выделены в Африке (вирус Ласса), а также широко распространены в Европе, Америке, Австралии и Азии (вирус лимфоцитарного хориоменингита). Несмотря на то, что аренавирусы содержат общие эпитопы, вирусы Нового и Старого Света различаются по поверхностным компонентам, строению кодирующих их генов, а также по видам грызунов, на которых они паразитируют.
Не получая поддержки от кровососущих членистоногих, аренавирусы передаются между зараженными животными через высохшие испражнения и респираторные секреты. Человек инфицируется при аэрозольном и алиментарном контактах с объектами, загрязненными выделениями инфицированных животных; заражение может происходить через поврежденную кожу. От человека человеку инфекция не передается.
Можно говорить о почти безвредной связи ареновирусов со своими основными хозяевами, причем природным резервуаром для каждого из них является обычно единственный вид грызуна. Это объясняет природную очаговость южно-американских лихорадок (естественные хозяева — грызуны Calomys callosus [вирус Мачупо], Calomys musculinus [вирус Хунин], Zygodontomys brevicauda [вирус Гуанарито]) и африканской лихорадки Ласса (естественный хозяин — многососковые крысы рода Mastomys [M. natalensis, M. еrythroleucus]). Что касается лимфоцитарного хориоменингита, то его повсеместное (кроме Африки) распространение тоже связано с экологией вируса. В качестве основного хозяина он использует серых домовых мышей (Mus domesticus, Mus musculus), которые до недавних пор служили единственным источником заражения людей. В последние годы описаны случаи лимфоцитарного хориоменингита, связанные с латентной инфекцией хомяков, в том числе из лабораторных колоний.
Аренавирусы имеют оболочечный вирион круглой или овальной формы, диаметром около 100 нм с колебаниями от 60 до 300 нм. Они содержат гранулярные частицы — клеточные рибосомы, давшие название вирусам (рис. 2).

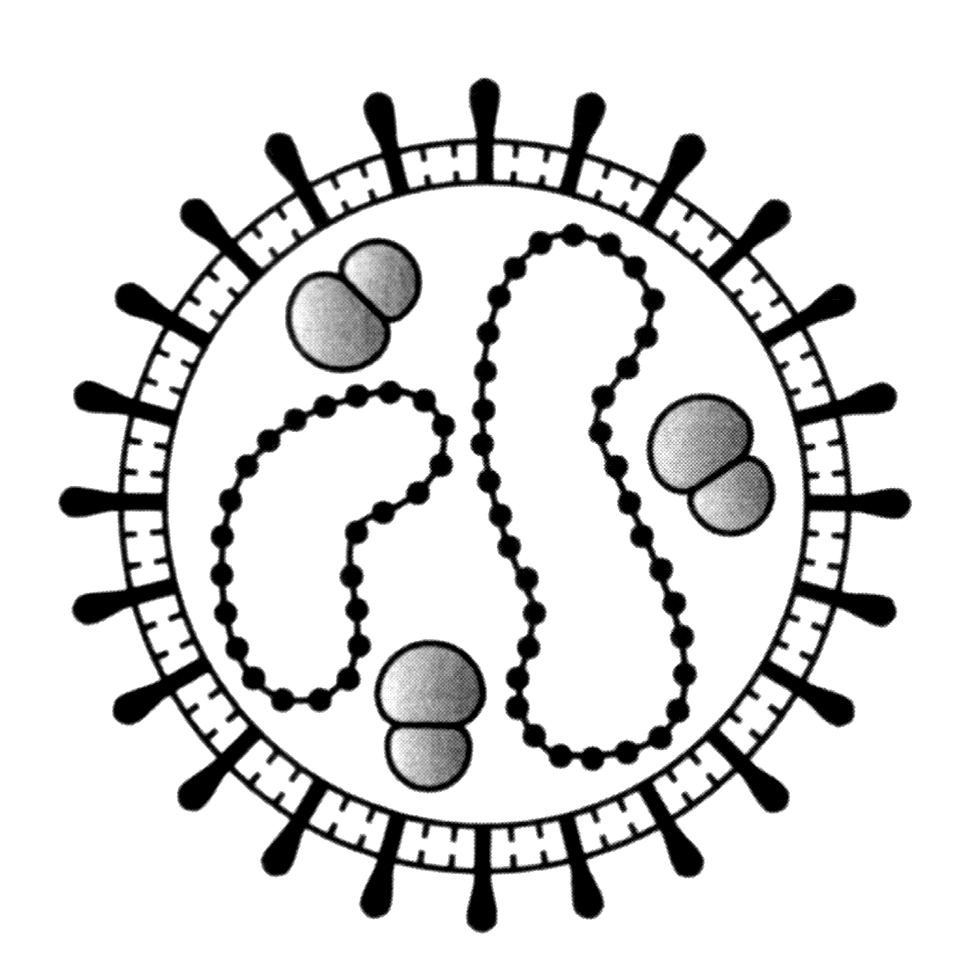
Роль рибосом не ясна, но они не определяют инфекционной силы вируса. Вирионная РНК состоит из двух циркулярных сегментов — L (англ. long) и S (англ. short) величиной 7,2 и 3,4 кбайт. Они находятся в спиралевидном комплексе с белком — N-протеином (англ. nucleocapsid). На поверхности вириона имеются шипы длиной около 10 нм, которые состоят из основания (гликопротеин G2) и головки (гликопротеин G1), связанной с G2 ионным взаимодействием. Белки N и G2 отличаются консерватизмом, определяя антигенные контакты между аренавирусами; для G1 характерна видовая и даже штаммовая специфичность. С вирусной частицей связаны также белки L (РНК-зависимая РНК-полимераза) и Z (функции неизвестны); их положение в вирионе не определено.
Репликация происходит в цитоплазме. РНК — односпиральна, отличается так называемой амбисенс-стратегией кодирования, т.е. ведет себя как минус- и плюс-РНК.
Поэтому хотя об аренавирусах принято говорить как о негативных РНК-вирусах, это неверно.
- Примерно половина S-сегмента ведет себя как минус-РНК, определяя синтез нуклеокапсидного (N) протеина.
- Другая часть является плюс-РНК, обеспечивая синтез предшественника поверхностных гликопротеинов.
L-сегмент вирионной РНК негативно кодирует L-белок, РНК-зависимую РНК-полимеразу, которая, в частности, определяет синтез небольшого (11 кДа) цинксвязывающего белка с неясными функциями.
Полипротеин после гликозилирования расщепляется на G1 и G2. Они встраиваются в плазматическую мембрану и служат местом взаимодействия с вирусным нуклеокапсидом. Обретая липидную оболочку (суперкапсид), вирус отпочковывается от клетки. G1 и G2 идут на построение вирусных шипов, предназначенных для взаимодействия с клеточными рецепторами, и служат мишенью для нейтрализующих антител.
Резервуаром для аренавирусов является хроническая инфекция грызунов. Заражение обычно проходит бесследно, причем лучшим вариантом является раннее инфицирование (плоды, новорожденные), когда организм отвечает иммунологической толерантностью на внедрение вируса. Возникает персистентная вирусемия, которая отражает недостаточный иммунный ответ у зараженных животных. Некоторые аренавирусы (например, вирус Ласса) используют механизм вертикальной передачи от матерей потомству. В других случаях раннее инфицирование достигается при сбрасывании в мочу вируса, что предполагает продолжение жизни аренавируса за счет заражения новых особей.
Человек является случайным хозяином аренавирусов. Заражение, как правило, не дает клинического результата, но иногда может иметь тяжелые, часто летальные последствия. О природноочаговом характере инфекций говорят названия основных аренавирусных болезней — Аргентинская (вирус Хунин), Боливийская (вирус Мачупо), Венесуэльская (вирус Гуанарито) латиноамериканские лихорадки, африканская лихорадка Ласса (вирус Ласса). Они отражают место первого описания болезней и их вирусов. Только лимфоцитарный хориоменингит избежал географической участи и распространен повсеместно через синантропных мышей.
По клинике большинство инфекций не выходит за рамки гриппоподобных заболеваний. После 10—14 дней инкубационного периода поднимается температура, возникают озноб, головная и мышечные боли, недомогание, боль в горле, артралгии. При тяжелых формах через неделю появляются симптомы геморрагического диатеза и поражений нервной системы (кожные и мукозальные петехии, кровотечения, рвота, диарея, дизурия, фотофобия, потеря слуха, иногда гепатит и др.). Повышение проницаемости капилляров ведет к оттоку плазмы в ткани, сгущению крови и шоку. Повреждение стволовых клеток костного мозга сопровождается лейкопенией и тромбоцитопенией (одна из причин геморрагического синдрома и кровотечений). В целом болезнь протекает благоприятно и заканчивается выздоровлением через 2—4 нед. Летальность — менее 2%.
Патогенез малопонятен. Гриппоподобные симптомы говорят о том, что аренавирусы, проникая через верхние дыхательные пути (контакт с высохшей мочой грызунов), вызывают повреждение респираторного эпителия. Размножение продолжается в регионарных лимфатических узлах, откуда вирусы попадают в кровь, где обнаруживаются в течение 3 нед после начала инфекции. Возбудитель присутствует в различных органах, но степень патологии не соответствует вирусологическим находкам. Попытки связать развитие болезни с иммунокомплексным воспалением оказались безрезультатными. Между тем, по аналогии с естественными хозяевами (грызунами) иммунологический конфликт должен играть важную роль в аренавирусной патологии. В опытах на мышах показано, что инфекция новорожденных ведет к длительной и безвредной персистенции вируса лимфоцитарного хориоменингита. Заражение взрослых особей вызывает острую смертельную нейроинфекцию, связанную с иммунными сдвигами, прежде всего в Т-клеточном звене иммунитета.
Учитывая контактную безвредность аренавирусов для клеток грызунов и человека, трудно допустить значение прямого цитопатического эффекта. Следует задуматься о медиаторах (тех же цитокинах), выделяемых зараженными клетками, и их участии в формировании дисфункций. В частности, мышей, зараженных летальной дозой вируса лимфоцитарного хориоменингита, удавалось спасти при обработке антителами к альфа/бета-интерферону. Это говорит о том, что эндогенные цитокины повышают патогенетическую активность аренавирусов, тем более что сами они мало чувствительны к действию интерферонов.
Элиминация аренавирусов у человека связана с нейтрализующими антителами, которые образуются чрезвычайно медленно, но, появившись, сохраняются долгие годы. Роль клеточного иммунитета в саногенезе при аренавирусных заболеваниях подлежит уточнению.
Клиническая диагностика аренавирусных инфекций (это относится ко всем арбо- и родентвирусным болезням) представляет большие трудности. Поэтому особое значение имеет специфическая диагностика, основанная на выявлении возбудителя и антивирусных антител. Материалом для выделения вируса служат кровь и спинномозговая жидкость больных, которые вводят в культуры клеток или интрацеребрально сосущим и только что отлученным от матери мышам и хомякам; при подозрении на лимфоцитарный хориоменингит и лихорадку Ласса используют взрослых мышей. О присутствии аренавирусов можно судить по наличию РНК в ПЦР с обратной транскрипцией. Для поиска вирусных антигенов применяют иммуноферментный тест. Наконец, возможно определение антивирусных антител. Из различных методов для этого лучше всего подходит реакция с энзиммечеными антителами.
Неспецифическая профилактика основана на борьбе с грызунами. Разработана живая аттенуированная вакцина из вируса Хунин, которая прошла успешное испытание на людях в странах, эндемичных по Аргентинской геморрагической лихорадке. Получена рекомбинантная вакцина для вируса Ласса. Она оказалась высоко протективной в опытах на зараженных обезьянах.
Вирус иммунодефицита человека
Его догнать пытался я, но он исчез,и снова появился, и вновь исчез…
- История открытия и происхождение.
- Вирион.
- Молекулярные и клеточные мишени.
- Взаимоотношения вируса с клеткой.
- Персистенция и агрессивность.
- Патогенез, патология, фазы инфекции.
- Синдром приобретенного иммунодефицита.
- Иммунитет: проигранная стратегия.
- Лабораторная диагностика.
- Принципы этиотропной терапии.
- Проблемы превентивной и стерилизующей вакцинации.
В 1981 г. Центром по контролю за заболева- емостью в США было сделано сообщение о том, что в течение восьми месяцев в Лос-Анжелесе зарегистрировано пять больных с редкой формой пневмонии, вызванной пневмоцистами — Pneumocystis cаrinii (сегодня их относят к грибам — см. «Медицинская микология»). Цифра впечатляла, так как за предыдущие десять лет было зарегистрировано всего два случая такой патологии. В связи с тем, что пневмоцисты не опасны для здоровых людей и проявляют агрессивность лишь на фоне ослабленного иммунитета, возникло подозрение о новом заболевании, которое спровоцировало иммунную недостаточность, осложненную пневмоцистозом. В отчете отмечалась еще одна подробность: все больные были молодыми гомосексуалистами.
Примерно тогда же были собраны данные о новых случаях саркомы Капоши. Ранее эту разновидность рака наблюдали у пожилых мужчин и лиц, принимавших иммунодепрессанты. На сей раз болели молодые гомосексуалисты. У некоторых из них саркома Капоши сочеталась с пневмоцистной пневмонией.
Вскоре появилось еще два сообщения — о повышении числа больных с хронической лимфаденопатией (стойким увеличением лимфатических узлов) и редкой формой злокачественной опухоли лимфоидной ткани, именуемой диффузной недифференцированной неходжкинской лимфомой. Оба наблюдения относились к гомосексуалистам и были расценены как результат поражения иммунной системы неизвестным агентом.
В 1982 г. была утверждена новая нозологическая единица — синдром приобретенного иммунодефицита, СПИД (англ. AIDS, от aquired immunodeficiency syndrome). Лабораторные анализы выявили главную аномалию у таких больных — резкое сокращение количества циркулирующих CD4 Т-лимфоцитов (тогда их называли Т4), центрального узла регуляции иммунного ответа.
Появились данные о заражении СПИДом через препараты крови (больные гемофилией) и парентеральные процедуры (наркоманы, пользующиеся общими шприцами). Стало известно о случаях СПИДа, связанных с гетеросексуальным и перинатальным заражением. Оказалось, что СПИД — лишь верхушка айсберга, основу которого составляют носители клинически латентной или малосимптомной инфекции.
Вслед за США СПИД был обнаружен в Центральной Африке и на Гаити. Болезнь быстро перекочевала в Европу, ознаменовав начало пандемии. По глобальным (скорее всего неполным) подсчетам в настоящее время заражено более 30000 млн. человек. Страны бывшего СССР включились в пандемический процесс позднее, но сегодня Россия и другие постсоветские республики быстро догоняют лидеров.
Возбудитель СПИДа открыт в 1983 г. Р. Галло (США) и Л. Монтанье (Франция). Он получил название вирус иммунодефицита человека, ВИЧ (англ. HIV, human immunodeficiency virus). Спустя два года был обнаружен еще один вирус, связанный с этиологией СПИДа. Он выделен от жителей Западной Африки, прибывших на лечение в Европу. Клиника СПИДа сочеталась у них с отрицательными серологическими реакциями на уже известный ВИЧ, который с этого момента стали называть ВИЧ-1. Новый тип вируса получил следующий порядковый номер, ВИЧ-2. Он близок ВИЧ-1, но менее болезнетворен. Распространение ВИЧ-2 ограничено странами Западной Африки.
Следы ВИЧ ведут к 1950-м гг. Именно к этому периоду относится обнаружение анти-ВИЧ антител в законсервированных сыворотках, полученных от жителей Африки. Африканский континент считается колыбелью ВИЧ, где он, пройдя многоэтапную эволюцию, созрел для выхода на мировую арену. В качестве стартового толчка для ВИЧ-1 предполагается заражение человека от шимпанзе. От них удалось выделить несколько штаммов вируса, генетически близких ВИЧ-1, в частности, по vpu-гену (см. ниже), которого нет у ВИЧ-2. ВИЧ-2 (или очень близкий к нему вирус) безболезненно заражает мангобеев и некоторые виды африканских обезьян. В то же время многие виды обезьян (например, макаки), сталкиваясь с ВИЧ-2 и не будучи его естественными хозяевами, болеют СПИДом. С этой точки зрения, ВИЧ-2-инфекцию можно считать зоонозом, тогда как ВИЧ-1 — антропоноз, распространенный только среди людей.
ВИЧ-1 остается лидером в изучении вирусов иммунодефицита, и все, о чем пойдет речь, относится прежде всего к нему. Впрочем, сходство ВИЧ-1 и ВИЧ-2 позволяет распространить большинство фактов на оба вируса. Поэтому мы будем пользоваться единым термином, ВИЧ, без разделения на типы.
ВИЧ принадлежит к семейству ретровирусов. Это РНК-вирусы с уникальным типом репродукции, которая в качестве обязательного этапа включает транскрипцию вирусного РНК-генома в комплементарную молекулу ДНК. Иными словами поток генетической информации временно получает обратное течение — от РНК к ДНК, вместо классического — от ДНК к РНК. Этот механизм, называемый обратной транскрипцией (англ. reverse transcription), послужил поводом для наименования семейства, Retroviridae.
ВИЧ включен в одно из подсемейств ретровирусов — медленные вирусы (лентивирусы). Все лентивирусы (их менее десятка) вызывают инфекции, имеющие общие признаки: длительный инкубационный период, бессимптомная или малосимптомная персистенция, мультиорганные поражения и неизбежно смертельный исход.
Лентивирусы человека представлены вирусами иммунодефицита, ВИЧ-1 и ВИЧ-2. Кроме человека, к ним восприимчивы только шимпанзе, но они заражаются на непродолжительное время без клинических последствий. Сходные вирусы животных представлены вирусами иммунодефицита обезьян и кошек. Главный резервуар одного из основных обезьяньих вирусов — африканские зеленые мартышки: их популяции заражены на 30—70%. Как уже говорилось, вирус не вызывает болезни у естественных хозяев, но может быть причиной СПИДа у других видов обезьян (в эксперименте им заражают макак). Кошачий вирус — единственный вирус животных, который, подобно ВИЧ, вызывает СПИД у своих естественных хозяев (кошек).
Вирион, субтипы и мишени
Устройство и внешний вид ВИЧ-вириона типичны для ретровирусов. Это частицы сферической формы, диаметром около 120 нм. В них хорошо контурируется нуклеокапсидная сердцевина (кор), окруженная наружной оболочкой (суперкапсидом). В отличие от других ретровирусов (в том числе ближайших соседей по таксономике — лентивирусов) сердцевина имеет коническую форму и расположена эксцентрично (рис. 1); у других ретровирусов сердцевина — правильной шаровидной формы и находится в центре вириона. Даже эти, чисто морфологические детали говорят об особой позиции ВИЧ в ретровирусном семействе.

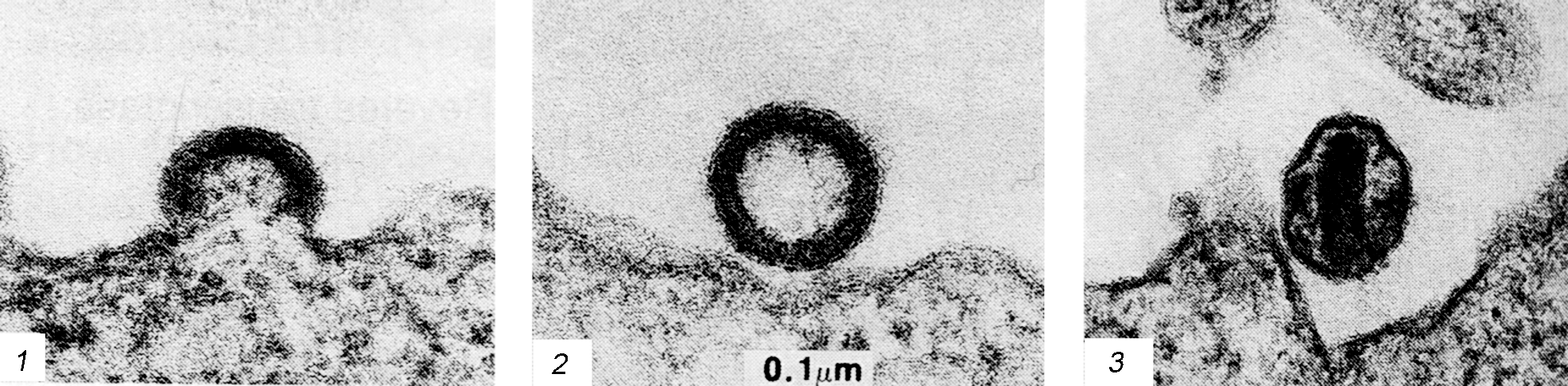
Принцип строения ВИЧ-вириона показан на рис. 2. Сердцевина содержит РНК-геном (представлен двумя идентичными молекулами РНК) и ферменты — обратную транскриптазу (р66/51), РНК-азу (р15), эндонуклеазу/интегразу (р32) и протеазу (р12). Все это находится внутри белкового чехла (капсида), построенного по типу кубической симметрии из белка р24. РНК стабилизирована двумя белками — р7 и р9. Суперкапсид образуется из плазматической мембраны, дополненной вирусными гликопротеинами (gp; от англ. glycoprotein). Их два — трансмембранный (gp41) и нековалентно связанный с ним gp120, выступающий в виде шипов. С внутренней стороны к суперкапсиду прилегает матриксный каркас, образованный белком р17.

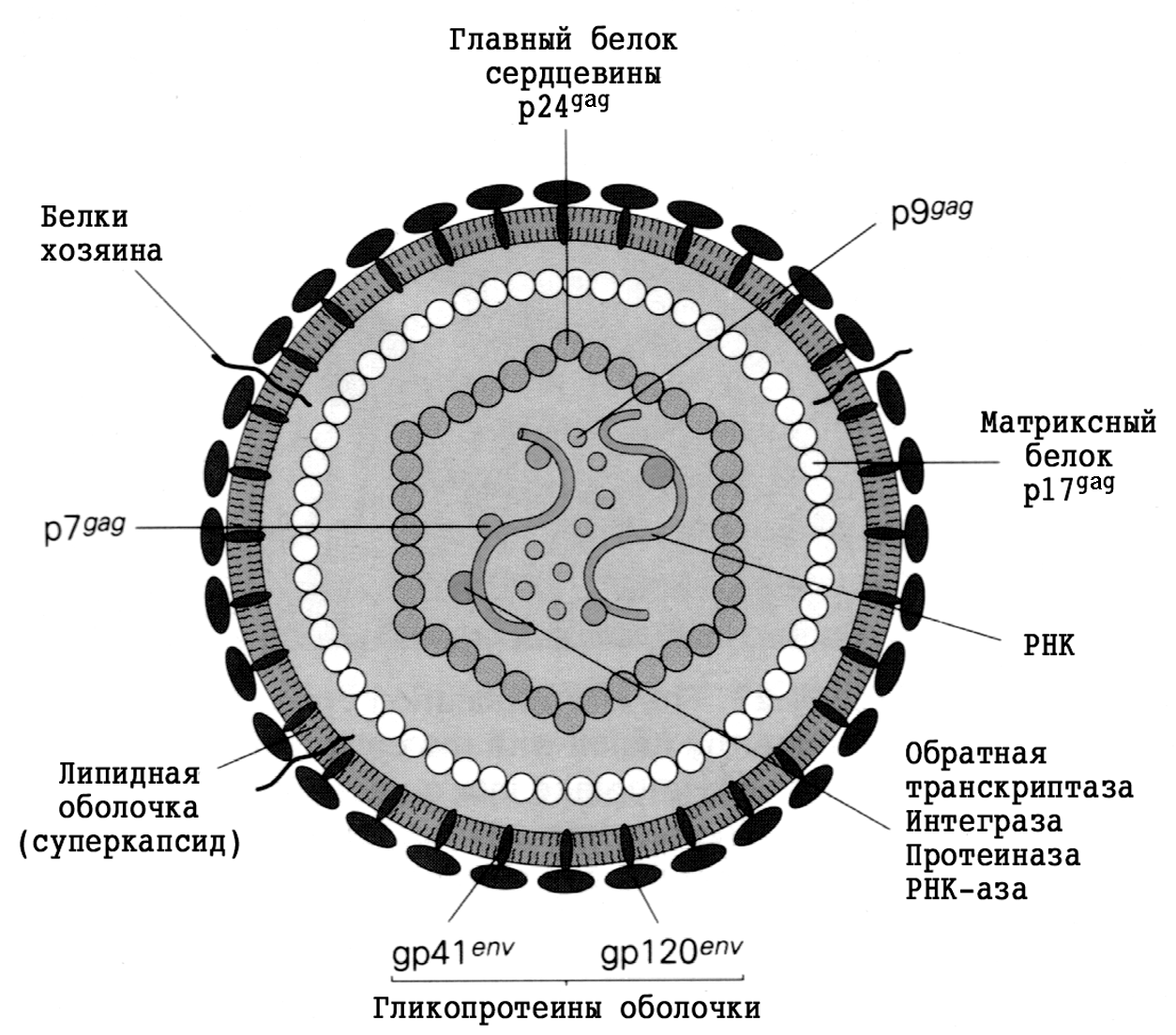
Образование вирионных (структурных) белков закодировано в трех генах. Продукт каждого из них представляет собой полипротеин-предшественник, который после расщепления (протеолиза) превращается в зрелые белки. Белки сердцевины и матриксный белок синтезируются в виде предшественника, закодированного в gag-гене (англ. group antigen). Вирионные ферменты возникают на основе продукта pol-гена (англ. polymerase). Суперкапсидные белки образуются при расщеплении сильно гликозилированного протеина на основе env-гена (англ. evelope). Протеолиз протекает в несколько этапов под действием вирусной и клеточных протеаз (см. ниже).
Генетический анализ ВИЧ-изолятов, полученных в различных регионах земного шара, утвердил представления о неоднородности ВИЧ. По особенностям нуклеотидного профиля env- и gag-генов предложено дифференцировать субтипы, или клады (греч. klados — ветвь) ВИЧ-1, которые объединены в три основные группы: М, N и О. Подавляющее большинство случаев ВИЧ-1 инфекции связано с вирусами М-группы (англ. major — главный). Им противопоставлена О-группа (англ. outliner — в стороне), которая включает небольшое число вариантов преимущественно из Западной Африки. N означает штаммы ВИЧ-1, не относящиеся к группам М и О.
М-группа содержит субтипы — от А до I. В странах Америки, Западной Европы, в Австралии и Японии преобладает субтип В (около 50%); вслед за ними идут субтипы А (16%), С, D и Е (по 10% каждого). Другие субтипы встречаются гораздо реже. В Африке доминируют субтипы А и С, в странах Юго-Восточной Азии чаще встречается субтип Е. Эпидемия в Индии вызвана субтипом С.
Первый случай СПИДа на территории Российской Федерации был вызван вирусом субтипа А. В период 1987—1990 гг. в России выделялись вирусы субтипов В, G и C. В настоящее время в России доминирует субтип А, распространяющийся среди наркоманов (более 75% случаев).
Доказана возможность сочетанного заражения разными кладами с возникновением мозаичных (рекомбинантных) вариантов ВИЧ-1. Они составляют около 10% всех ВИЧ-геномов с известной нуклеотидной последовательностью. В настоящее время известны все возможные сочетания, по крайней мере, для субтипов А—D, F—H.
Развитие ВИЧ-инфекции — многофазный и многофакторный процесс, в основе которого лежит прогрессирующее поражение иммунокомпетентных клеток. Главной мишенью для вируса служит хелперная субпопуляция Т-лимфоцитов. Это объясняется тем, что Т-хелперы несут на своей поверхности рецептор CD4, который связывается с одним из поверхностных ВИЧ-белков — gp120. CD4 Т-лимфоциты обеспечивают более 90% репликативного выхода вируса.
Вторыми по значимости являются мононуклеарные фагоциты: около 40% моноцитов крови и тканевых макрофагов содержат CD4. Известны и другие категории CD4-позитивных клеток (дендритные клетки, естественные киллеры, нейроны), но этот признак выражен у них гораздо слабее и менее постоянно.
Есть клетки, которые не имеют CD4 рецепторов, но тем не менее чувствительны к ВИЧ. К ним относятся М-клетки слизистой прямой кишки и сперматозоиды. Это означает, что по крайней мере часть вирусных клонов воспринимается независимо от CD4 за счет дополнительных клеточных структур. Установлено, что ВИЧ взаимодействует с маннозосвязывающим белком макрофагов и дендритных клеток, галактозилцерамидом нейроглии, рецепторами, сопряженными с G-белками. Они реагируют все с тем же gp120, но сайты связывания не совпадают между собой и с лигандом для CD4. Известны и примеры наведенной (опосредованной) ВИЧ-рецепции за счет связывания вирионов, покрытых антителами и С3-фактором комплемента.
Чтобы попасть в клетку, вирусу необходим второй суперкапсидный белок (gp41): он обеспечивает слияние клеточной и вирусной мембран благодаря связыванию с F-пептидом клеток (от англ. fusion — слияние). Синтезирован пептид, который после связывания с gp41 препятствует репликации вируса в чувствительных клетках. Это делает его одним из кандидатов для включения в комплексную анти-ВИЧ-терапию.
Но главными кофакторами ВИЧ-заражения служат два хемокиновых рецептора, СХСR4 и CCR5. Вместе с CD4 они взаимодействуют с gp120, раскрывая функцию слияния gp41. Хемокины, т.е. естественные лиганды для CXCR4 и CCR5, блокируют этот процесс, задерживая размножение ВИЧ в лимфоцитах. Врожденный дефицит по CXCR4 и CCR5 (не опасный для его обладателей) рассматривается как фактор, предохраняющий от ВИЧ-инфекции.
Небезразличным считается и присутствие в ВИЧ-оболочке белков клетки-хозяина. Некоторые из них (например, HLA-2, CD54, CD11a), возможно, вносят вклад в особенности реакций на внедрение вируса — от активации до апоптоза клеток.
Жизненный цикл
Репликация ВИЧ (рис. 3) не отличается от других ретровирусов, но находится под более сложным контролем, который включает факторы позитивной и негативной регуляции. Их активность определяет соотношение между латенцией вируса и продуктивной (гибельной для клетки) инфекцией.
Вирус проникает в клетку путем рецепторзависимого эндоцитоза. После слияния плазматической и суперкапсидной мембран сердцевина вириона оказывается в цитоплазме. Далее с помощью обратной транскриптазы производится синтез комплементарной ДНК. Вирусная РНК-аза разрушает использованную РНК-матрицу, а обратная транскриптаза, действуя как обычная ДНК-полимераза, достраивает вторую ДНК-нить. Это происходит быстро: уже через 2 ч после инфицирования в цитоплазме и ядре обнаруживаются вирусные ДНК.

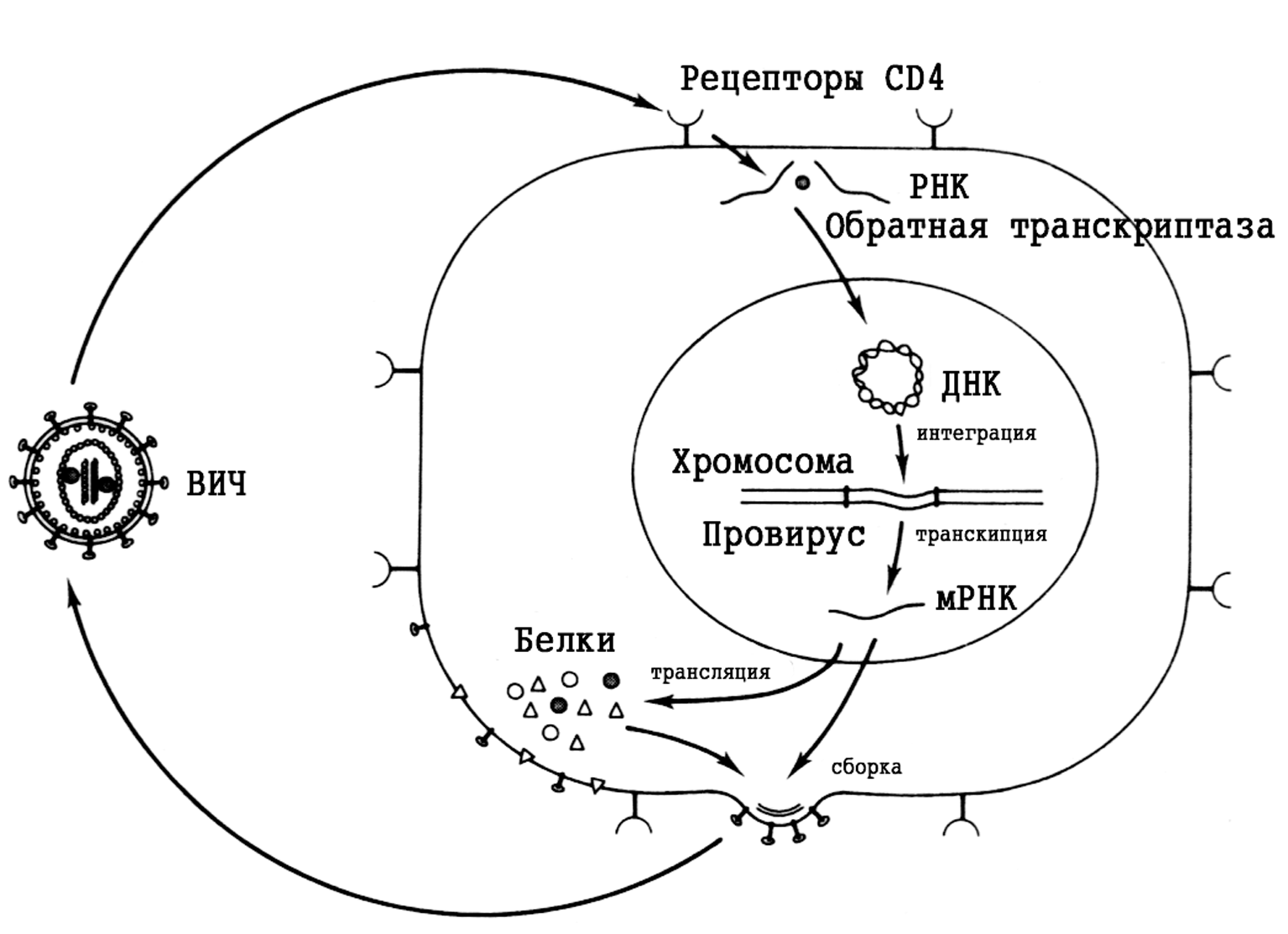
Синтез ДНК сопровождается важным для вируса событием: концевые участки геномной РНК копируются дважды, образуя так называемые длинные концевые повторы, LTR (от англ. long terminal repeat) — около 630 пар нуклеотидов на каждом из концов ДНК. Они необходимы для включения вирусной ДНК в состав хромосом и для дальнейшей регуляции вирусных генов.
Готовая ДНК (она временно обретает замкнутую кольцевидную форму, предохраняющую от разрушения клеточными нуклеазами) перемещается в ядро, где включается в хромосомы. Этот процесс, называемый интеграцией, реализуется вирионным ферментом — интегразой. Действуя как эндонуклеаза, она разрезает в определенных местах клеточную ДНК, соединяя рассеченные концы с LTR вируса. С этого момента генетический материал ВИЧ называется провирусом. Он ведет себя как обычный клеточный ген, переходя при делении в дочерние клетки и воспринимая сигналы, контролирующие транскрипцию.
Перед вирусом стоит альтернатива. Он может надолго выйти из игры, ничем о себе не заявляя, либо перейти в продуктивную фазу, занявшись воспроизведением собственного потомства. В этом случае вирусная ДНК транскрибируется клеточной РНК-полимеразой, направляемой сигналами с LTR. Часть синтезированных РНК-молекул используется как мРНК, другие (полные по длине транскрипты) служат репликативными РНК для новых вирусных частиц. Сборка вирионов завершается формированием наружной оболочки. Это происходит при почковании через участки плазматической мембраны с встроенными суперкапсидными белками (cм. рис. 1).
В ходе репликативного процесса ВИЧ синтезирует три полипротеина (Gag, Pol и Env), которые служат основой для получения структурных белков. Вирионная протеаза (она аутокаталитически вычленяется из Pol-полипротеина) доводит до кондиции белки-предшественники сердцевины (Gag) и ферментного комплекса (Pol) (рис. 4). Образование суперкапсидных белков (gp120 и gp41) обеспечивается из Evp-предшественника клеточной протеазой.

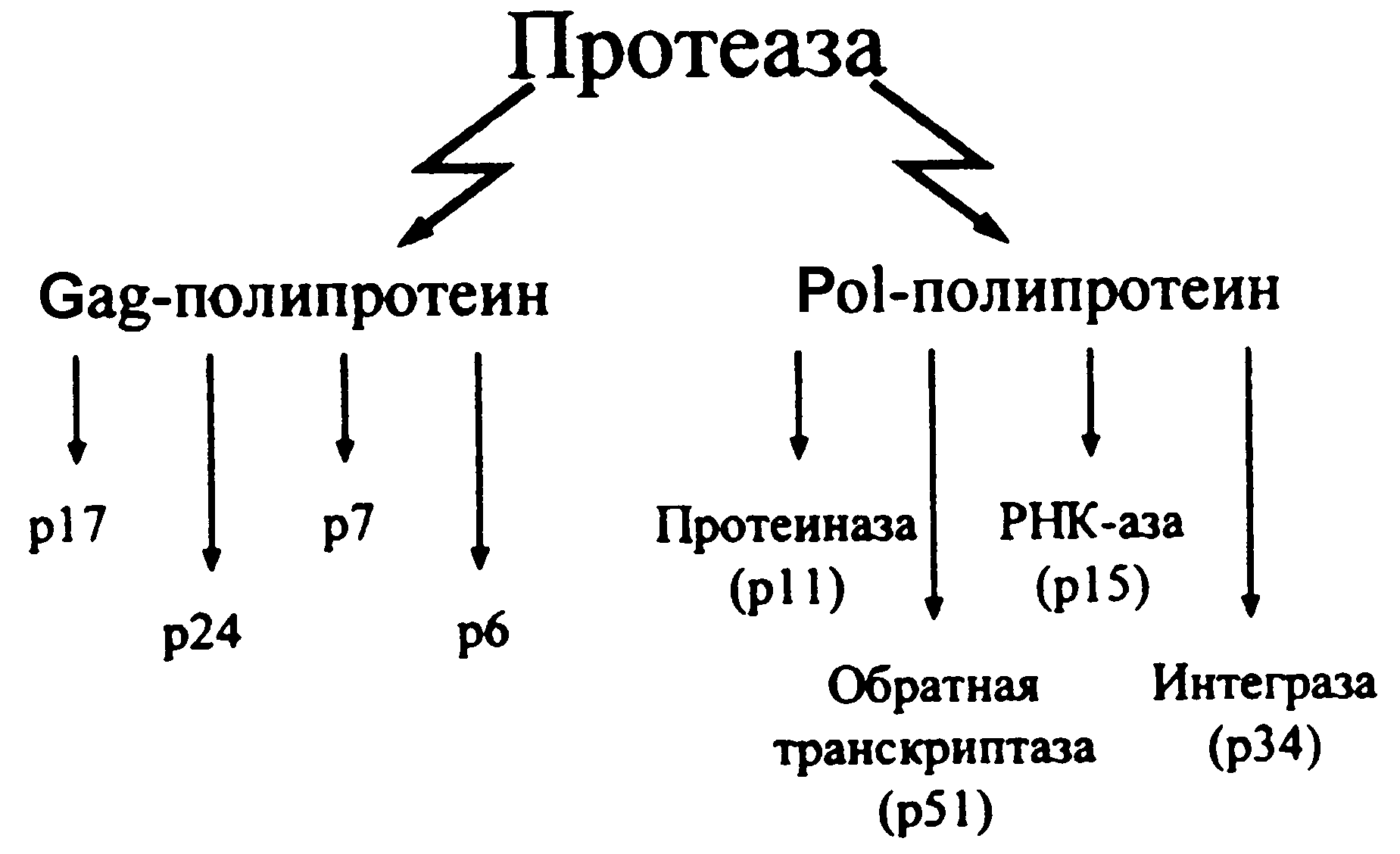
Чтобы поддерживать репродукцию вируса, клетка должна быть метаболически активной. У лимфоцитов этому соответствует фаза пролиферации, возбуждаемая антигенами или цитокинами. Это непрерывный и неизбежный процесс, который периодически усиливается или ослабевает. На уровне организма ВИЧ-инфекция никогда не достигает полной латенции, и признаки активной репликации в виде более или менее выраженной вирусемии всегда налицо. Это поддерживает непрерывный и исключительно интенсивный кругооборот вирионов: обновление циркулирующего ВИЧ-пула происходит каждые два часа.
Для ВИЧ характерно наличие генов, кодирующих неструктурные (регуляторные) белки (пептиды) — tat, rev, vpr, nef, vif, vpu и vpr. Ими обладают только лентивирусы, причем по разнообразию регуляторных генов ВИЧ превосходят другие медленные вирусы. Первыми проявляют себя tat-, rev- и nef-гены. Пептид Таt (мол. масса 14 кДа; англ. trans-activation) является универсальным активатором транскрипции ВИЧ-генов. Под его влиянием она усиливается в 50—1000 раз, причем стимулирующий эффект распространяется на синтез всех вирусных белков — структурных и неструктурных.
Rev-ген (англ. regulator of virus) действует более осмысленно. C появлением его продукта (пептид с мол. массой 19 кДа) доминирует синтез вирионных белков. При относительном балансе между Тat- и Rev-пептидами репликация вируса ограничена, позволяя ему воспроизводиться годами без грубого повреждения клеток. Такой тип регуляции логично рассматривать как выгодное для вируса приспособление, нацеленное на адаптацию к долгоживущим хозяевам.
Nef-пептид (англ. negative-regulatory factor; мол. масса 27 кДа) вызывает негативную регуляцию ВИЧ-инфекции. Он ослабляет экспрессию всех генов, подавляя Тat-зависимую активацию провируса. Однако этот эффект много слабее Тat-зависимой экспрессии генов.
Синтез Vif-, Vpu- и Vpr-пептидов вместе с продукцией вирионных белков относится к поздней фазе ВИЧ-репликации. Наиболее очевидна феноменология Vif-эффекта (англ. virion infectivity factor; мол. масса 24 кДа): он усиливает способность свободных вирионов инфицировать соседние клетки. Полагают, что Vif-фактор каким-то образом следит за сборкой сердцевины, от которой зависит активность вирусного генома в зараженных клетках.
Ген vpu (англ. virus protein U) уникален для ВИЧ-1. Его нет у других лентивирусов, а у ВИЧ-2 ему соответствует vpx-ген. Cчитают, что Vpu-пептид (мол. масса 16 кДа), усиливает трансмембранный экспорт (почкование) вирусных частиц. При vpu-мутациях появляются уродливые вирионы (с несколькими сердцевинами), которые не выделяются во внешнюю среду, а отпочковываются во внутриклеточные вакуоли. Кроме того, Vpu-фактор снижает экспрессию CD4-рецепторов на Т-лимфоцитах, что нарушает взаимодействие иммунокомпетентных клеток.
Продукт vpr-гена (англ. virus protein regulator; мол. масса 15 кДа) обнаружен в зрелых вирионах. Среди его возможных функций назывались 1) регуляция обратной транскрипции, 2) стабилизация РНК и ДНК-структур, 3) транспорт ДНК в ядро, 4) регуляция интегративного процесса, 5) стартовая активация вирусных генов. Не исключено, что Vpr-пептид более важен для взаимодействия вируса с макрофагами, чем с лимфоцитами.
Рассуждая о регуляторных генах, следует помнить, что они были бы бесплодны, если бы вирус не умел управлять их транскрипцией. Эту функцию выполняют LTR, фланкирующие провирус (см. выше). Их можно сравнить с пультом управления, насыщенным генетическими кнопками. Такую роль выполняют нуклеотидные последовательности, которые специфически связываются с регуляторными ВИЧ-пептидами и клеточными регуляторами транскрипции, усиливая или блокируя экспрессию вирусных генов.
Заражение
Единственным естественным источником ВИЧ (ВИЧ-1) является человек. Зараженные люди опасны на любой из стадий инфекции за исключением короткого промежутка времени от заражения до выхода вируса в кровь. Следует помнить, что вирусемия опережает сероконверсию (появление в крови анти-ВИЧ-антител), и в таких случаях суждение о «незаразности», основанное на негативных серологических тестах, будет ложным. Помимо крови, ВИЧ можно выделить из семенной жидкости, секрета цервикального канала, материнского молока, спинномозговой жидкости, слез, слюны, мочи. Эпидемически значимые концентрации вируса возникают только в крови, сперме, секрете шейки матки и (редко) в грудном молоке.
Обычный путь распространения вируса — половые контакты, как вагинальные, так и анальные. Опасность последних гораздо выше, так как однослойный эпителий прямой кишки намного ранимее многослойного эпителия влагалища, и вирус получает больше шансов для проникновения через микротравмы слизистой оболочки. Допускается и возможность заражения ректальных эпителиоцитов с образованием в них первичного резервуара ВИЧ-инфекции. Среди гетеросексуальных партнеров вероятность заражения колеблется в широких пределах — от 10 до 70%. Например, жены ВИЧ-позитивных гемофиликов часто не заражаются на протяжении нескольких лет регулярных половых контактов. В других случаях передача вируса происходит после одного-двух сеансов.
Известны примеры незаражения половых партнеров (гомосексуалистов и проституток), имевших неоднократные половые связи с ВИЧ-позитивными партнерами без всяких мер защиты. Можно размышлять о генетической природе резистентности к инфекции (например, гомозиготные мутации по CCR5-гену), повышенной вирулентности вирусных клонов, возникающих в ходе инфекционного процесса, о кофакторах передачи, предрасполагающих к заражению (травмированность слизистых гениталий и прямой кишки инфекционной и неинфекционной природы), о неодинаковой контагиозности ВИЧ-носителей в разные фазы инфекции в связи с колебаниями репликативного цикла.
Другой путь — передача вируса через зараженную кровь. Среди первых пострадавших от ВИЧ были больные гемофилией, получавшие постоянные вливания препаратов факторов VIII и IX. С введением жесткого контроля за донорами и разработкой способов стерилизации препаратов их опасность сведена к минимуму. Недавно стали доступны рекомбинантные антигемофильные факторы, которые полностью исключают возможность заражения ВИЧ и вирусами парентеральных гепатитов.
То же самое можно сказать и о медицинских процедурах. СПИД-трагедии, спровоцированные небрежным отношением к инъекционным операциям, известны медицинской практике, но стандартные меры предосторожности фактически исключают такую угрозу. Риск заразиться от случайного укола иглой, использованной для взятия крови у ВИЧ-инфицированных больных, не превышает 0,5%, а вероятность заражения при попадании инфицированного материала на слизистые оболочки и неповрежденную кожу еще ниже. Контакт с зараженной кровью, который легко предупредить в медицинских учреждениях, трудно устранить во внебольничных условиях. Не случайно ВИЧ-инфекция получила распространение среди наркоманов, пользующихся общими иглами и шприцами для введения наркотиков.
Драматична перинатальная ВИЧ-инфекция. Вирус проникает от матери к плоду/новорожденному трансплацентарным путем, во время родов (прохождение через инфицированный родовой канал) и, гораздо реже, с грудным молоком. По неизвестным причинам 60—80% детей, родившихся от ВИЧ-инфицированных женщин, лишены вируса. Но если предыдущий ребенок был инфицирован, риск заражения следующих детей возрастает. В отличие от взрослого варианта перинатальная ВИЧ-инфекция обычно быстро прогрессирует, завершаясь фатальным СПИДом в течение двух лет.
Вирус не передается при обычных бытовых контактах, через предметы обихода, аэрогенным путем. Это установлено многочисленными наблюдениями за членами семей ВИЧ-инфицированных и лицами, находившимися с ними в близком контакте. Безуспешными оказались попытки взвалить ответственность за заражение на кровососущих насекомых — комаров, вшей, постельных клопов и пр. Они не могут служить биологическим вектором ВИЧ, так как вирус не находит в них условий для размножения.
Ведутся споры о том, как ВИЧ попадает в организм — в виде свободных вирионов или внутри клеток, содержащих провирус и/или незрелые вирионы. Скорее всего, он использует оба способа. Внутриклеточный вирус недосягаем для антител и защититься от него путем вакцинации непросто, если вообще возможно.
Неодинакова и судьба вируса при разных способах заражения. Если ВИЧ поступает в кровь, его ожидает прямой маршрут в лимфоидную ткань, где он находит свои мишени, закрепляется в них и приступает к реализации своей агрессивности. При заражении через слизистые оболочки (половой путь) дело обстоит сложнее. В этом случае часть вирионов доставляется в лимфатические узлы дендритными клетками. ВИЧ использует такой прием, как дополнительный (а, возможно, и основной) механизм транспортировки к своим главным мишеням. Вирусные частицы, фиксированные на отростках дендритных клеток, доставляются в Т-зависимые зоны регионарных лимфатических узлов, где переключаются на Т-хелперы. Поэтому аккумуляция дендроцитов в зоне воспаления (например, при сопутствующих заболеваниях гениталий) может быть важным кофактором заражения ВИЧ. Дендритные клетки играют роль и в дальнейших событиях. Они косвенно поддерживают репликацию ВИЧ, провоцируя оживление провируса в активируемых Т-хелперах.
Изменчивость вируса
Гетерогенность, свойственная любым микробным популяциям, выражена у ВИЧ необыкновенно ярко. Фенотипическая вариабельность ВИЧ отражает неоднородность его генофонда: все ВИЧ-изоляты в большей или меньшей степени различаются по структуре (нуклеотидной последовательности) РНК. Отличаются не только штаммы от разных людей, но и клоны, выделяемые по ходу инфекционного процесса.
Высокая мутабельность генома характерна для всех ретровирусов. В этом отношении они превосходят другие РНК-вирусы, которые, в свою очередь, более изменчивы, чем ДНК-вирусы. Неспособность клетки исправлять ошибки вирусных РНК-зависимых полимераз (в данном случае РНК-зависимой ДНК-полимеразы) дополняется ошибками при интеграции и сплайсинге мРНК. Имеет значение и диплоидность ретровирусного генома, которая облегчает рекомбинацию между гетерозиготными нитями РНК. В целом на каждый репликативный цикл у ВИЧ приходится не менее десятка мутаций, а вариабельность его генома на несколько порядков выше, чем у вируса гриппа А, признанного лидера антигенных перестроек.
Для ретровирусов используется понятие квазивидов, подразумевающее, что уже первое потомство отличается от родителей (лат. quasi — почти, как будто). Это означает, что ВИЧ существует в виде популяций, состоящих из множества такого рода вариантов. Даже при бессимптомной инфекции ВИЧ подвергается непрерывной репликации, создавая мобильное потомство, готовое противостоять экологическому прессу хозяина. Особенно демонстративны эпитопные перестройки главного поверхностного антигена, gp120. В его структуре кроме относительно стабильных (консервативных) участков имеется несколько гипервариабельных доменов, подверженных высокой изменчивости. На этой основе формируются варианты ВИЧ, которым не страшны антитела и цитотоксические CD8 Т-лимфоциты, индуцированные их прародителями. Выживание таких ускользающих мутантов (англ. escape mutants) — один из главных механизмов ВИЧ-персистенции и камень преткновения на пути создания вакцины.
Альтернативными по цитопатичности являются разновидности вируса, вызывающие и не вызывающие образование синцития, т.е. многоядерных нежизнеспособных клеток. Это происходит в результате gp120-зависимого слияния мембран зараженных Т-лимфоцитов и соседних клеток. Мутационное замещение всего несколько аминокислот в одном из гипервариабельных доменов gp120 лишают вирус этого признака.
Различные ВИЧ-клоны с неодинаковой охотой размножаются в моноцитарно-макрофагальных и лимфоидных линиях клеток, хуже или лучше инфицируют дендроциты, клетки нервной ткани, эпителиоциты кишечника. Описаны и совсем необычные ВИЧ-варианты, которые реплицируются в эндотелиальных клетках, миоцитах и др. Штаммы, выделяемые на поздних стадиях болезни, лучше размножаются в лимфоцитах и инфицируют более широкий круг клеток. Это говорит об их повышенной агрессивности, обусловленной селекцией вирусных клонов в процессе длительного сосуществования с хозяином. Можно отметить, например, смену ВИЧ-клонов, использующих хемокиновые рецепторы. Почти все штаммы, которые начинают инфекцию, связываются только с CCR5. Позже появляются клоны, реагирующие с СXCR4. Это расширяет их вирулентность, позволяя атаковать циркулирующие CD4 Т-лимфоциты, которые в своем большинстве лишены CCR5 и поэтому не чувствительны к СCR5-клонам.
Патогенез и иммунология вич-инфекции
Обязательным условием болезнетворности ВИЧ является пожизненная персистенция возбудителя в организме человека. В ее основе лежит интегративная вирогения, т.е. включение вирусного генома в хромосомный аппарат клетки-хозяина. Но в отличие от других вирусов, склонных к латенции, интеграция ВИЧ непрерывно сочетается с его репликацией. Это делает персистенцию агрессивной, определяя патогенез ВИЧ-инфекции. Начавшись вскоре после заражения, репликативный процесс (об его интенсивности судят по вирусемии) протекает безостановочно с двумя пиками, в начале и в конце заболевания. Купирование первой волны репликации совпадает с появлением в крови анти-ВИЧ-антител, заключительный всплеск отражает финальный кризис иммунной системы. С вирусологической точки зрения у ВИЧ нет латентной фазы. После более или менее длительного периода стабилизации конфликт почти неизбежно завершается быстрой и фатальной развязкой.
Пройдя входные ворота, ВИЧ оседает в лимфатических узлах, приступая к воспроизведению потомства. Вскоре вирус обнаруживается в крови и, пользуясь отсутствием нейтрализующих антител, беспрепятственно распространяется по организму, поражая лимфоидную ткань, центральную нервную систему, эпителиоциты кишечника и другие ткани. Лимфатические узлы остаются главным резервуаром вируса и практически единственным источником новых вирионов. Это понятно, так как здесь собрана основная масса CD4 Т-лимфоцитов, готовых поддержать размножение вируса.
Сопряжение между репликацией вируса и хронической активацией иммунной системы по сути и есть та основа, на которой эволюционирует ВИЧ-патология. Поэтому агрессивность ВИЧ-инфекции (она определяет продолжительность инкубационного периода) во многом зависит от антигенных и других стимулирующих нагрузок, испытываемых CD4 Т-лимфоцитами. Это так называемые кофакторы ускорения, приближающие терминальную фазу заболевания. Потенцирующее действие оказывают сопутствующие инфекции, особенно если их возбудители привносят с собой поликлональные активаторы Т-лимфоцитов (суперантигены). Ускоряющий эффект посторонних вирусов может быть связан с трансактивацией ВИЧ-провируса. Таким эффектом обладает цитомегаловирус — один из наиболее вероятных кофакторов ВИЧ-инфекции.
Скорость ВИЧ-опустошения Т-хелперной популяции сопряжена с HLA-фенотипом (она выше у носителей HLA-A1, A5, A11, A23, B8, DR2, DR5 и ряда других HLA-аллелей), а устойчивость к заражению повышается при дефектах ВИЧ-корецепторов. Это показано для хемокинового рецептора CCR5. Установлено, что мутации по гену ССR5 служат причиной повышенной устойчивости к СПИДу около 25% населения северо-западных областей России.
Если на ранних этапах ВИЧ-инфекции СD4 Т-лимфоциты — почти единственная мишень для вируса, то по мере накопления мутантов (квазивидов) спектр клеточных мишеней расширяется. Т-хелперы остаются резервуаром продуктивной инфекции, но персистенцию вируса поддерживают и другие клетки, в первую очередь макрофаги. Они формируют второй по значимости резервуар вируса, оправдывая титул «второй ВИЧ-инфекции». Это не должно удивлять, так как и другие лентивирусы охотно заражают мононуклеарные фагоциты. Более того, патогенез ретровирусных инфекций у животных во многом опосредован через этот механизм.
Взаимоотношения ВИЧ с макрофагами складываются иначе, чем с Т-лимфоцитами. Они слабо поддерживают репликацию вируса, а главное не погибают от нее. Возможно, это связано с тем, что, закончив дифференцировку, макрофаги не пролиферируют (не размножаются). Поэтому вирус не получает в них полноценного набора кофакторов транскрипции, который имеется в активированных Т-лимфоцитах. Но именно это делает мононуклеарные фагоциты едва ли не главным хранилищем латентного вируса.
Не вызывая прямого повреждения, ВИЧ тем не менее заметно влияет на функции макрофагов. В них усиливается продукция цитокинов (в том числе активаторов Т-лимфоцитов) и снижается способность к представлению антигенов. Это чревато негативными последствиями. В первом случае возрастает опасность цитокининдуцированной репликации ВИЧ в CD4 Т-клетках, во втором — снижается и без того ослабленный потенциал иммунной системы. Есть основания полагать, что функциональные перекосы моноцитов/макрофагов играют ведущую роль в нейропатогенезе и, возможно, других нелимфоцитарных проявлениях ВИЧ-инфекции (см. ниже). Наконец, макрофаги, содержащие ВИЧ, хуже выполняют свои прямые обязанности, связанные с фагоцитозом. Это логично вписывается в патогенез иммунодефицитного ВИЧ-синдрома.
Агрессивность ВИЧ-персистенции во многом зависит от изменчивости вирионных антигенов, которая обеспечивает высокую устойчивость вируса к эффекторам иммунитета. В нейтрализации свободных вирионов почти все зависит от антител против оболочечных гликопротеинов, прежде всего gp120. Обновление поверхностных структур достигается благодаря высокой мутабельности env-гена, кодирующего суперкапсидные белки вириона, прежде всего вариабельные участки gp120. Но вирус укрывает и часть консервативных сайтов, экранируя их гепарвариабельными структурами. Наконец, чувствительность к антителам ослабляют клеточные белки, которые включаются в состав вирусной оболочки и прикрывают ее собственные структуры. Все это ведет к тому, что лишь немногие из антител обладают вируснейтрализующими свойствами, т.е. способны инактивировать инфекционный потенциал вируса.
В ощутимых титрах антитела появляются через несколько недель (1—3 мес) после снижения стартовой вирусемии. ВИЧ способен обезоруживать и нейтрализующие антитела. Это происходит благодаря их перехвату свободными молекулами gp120, которые сбрасываются с клеток при репликации вируса. Около половины gp120 попадают в кровь в растворимой форме.
Чтобы активно персистировать (т.е. непрерывно размножаться) вирус должен преодолеть еще одно препятствие. Синтез вирусных белков сопровождается «выныриванием» вирусных пептидов на поверхность зараженной клетки в составе молекул HLA. Комплексы HLA-1+ пептид распознаются цитотоксическими CD8 Т-лимфоцитами (ЦТЛ), которые уничтожают клетки, меченные вирусными антигенами. Содержание анти-ВИЧ ЦТЛ заметно опережает динамику антител и коррелирует с резким падением уровня ВИЧ-вирусемии, демонстрируя высокую эффективность данного механизма. Эпитопы, индуцирующие ЦТЛ, изучены хуже, чем В-эпитопы, но по крайней мере часть из них принадлежит вариабельным участкам gp120. Это означает, что они тоже втягиваются в мутагенез, периодически исчезая из поля зрения преформированных ЦТЛ. Удачное начало не завершается полной блокадой ВИЧ-репликации.
Так или иначе, но ВИЧ успешно выживает, несмотря на грозный арсенал нацеленных на него эффекторов иммунитета. Ликвидация острой вирусемии — лишь тактическая победа хозяина, стратегия остается за вирусом. Переводя конфликт в долгосрочную форму, он получает возможность реализовать свой патогенетический потенциал, побеждая в этой гонке с преследованием. Агрессивность ВИЧ-персистенции определяется поражением стратегически важного объекта — иммунокомпетентных клеток. С самого начала вирус втягивает их в порочный круг, заставляя работать на себя и тратить силы на ложные мишени. Это дезорганизует лимфоидную ткань, обезоруживая хозяина не только против ВИЧ, но и против других инфекций.
Число циркулирующих CD4 Т-клеток снижается уже на этапе первичной ВИЧ-вирусемии. Цитотоксические Т-лимфоциты и антитела приостанавливают этот процесс, но полного восстановления не происходит. После относительной компенсации (она продолжается несколько месяцев) количество Т-хелперов неуклонно сокращается и в терминальной фазе падает почти до нуля (100 клеток/мкл и ниже; в норме более 1000 клеток/мкл). Коэфициент СD4/CD8 в крови спускается ниже 1,0 (в норме 2,0 и выше).
Казалось бы, все очевидно: гибнут клетки, в которых размножается вирус. Но эта логика для ВИЧ слишком проста. На любом из этапов ВИЧ-инфекции заражается не более 5% Т-хелперов и поэтому даже при высокой скорости кругооборота вируса и реинфицирования новых мишеней таким путем уничтожается сравнительно небольшое количество клеток. Деструктивные ресурсы ВИЧ реализуются через дополнительные каналы. Прокомментируем некоторые из них.
Подобно другим оболочечным вирусам, ВИЧ индуцирует слияние клеток с образованием нежизнеспособных поликарионов. Это происходит в том случае, если вирусные белки (gp120), встроенные в плазматическую мембрану инфицированного Т-лимфоцита, связываются с рецепторами (CD4) соседних клеток (рис. 5). Единственная зараженная клетка способна втянуть в реакцию до 50 свободных от вируса соседей, что существенно расширяет масштаб повреждения. Мутационное замещение всего нескольких аминокислот в СD4-связывающем домене gp120 лишают вирус этого признака.

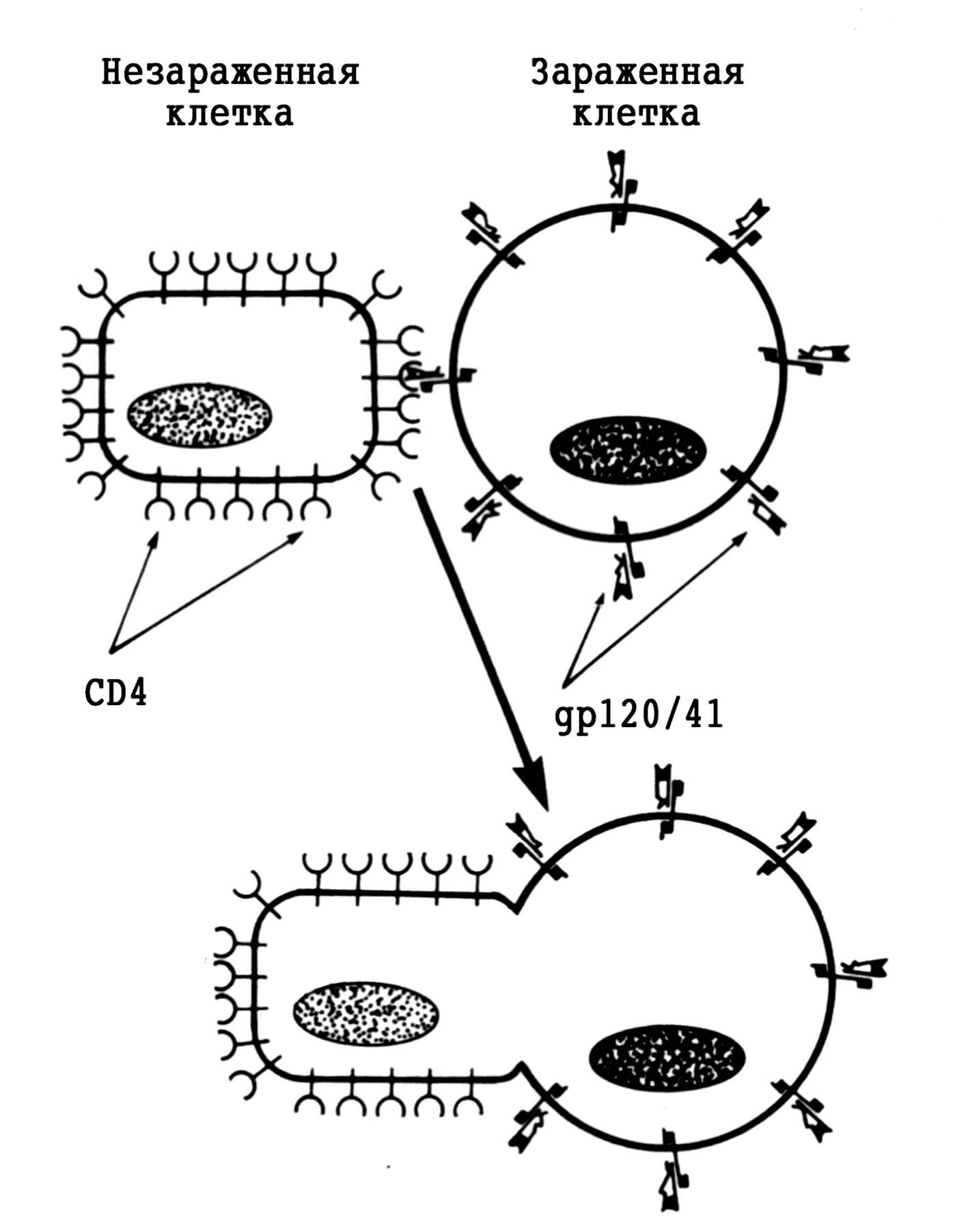
При репликации многие gp120 не включаются в вирионы, а сбрасываются в окружающую среду. Связываясь с CD4-рецепторами соседних клеток, свободный gp120 превращает их в потенциальную мишень для анти-ВИЧ-антител. Это предрасполагает к комплементзависимому цитолизу или к повреждению агрессивными клетками, несущими рецепторы для Fc-фрагмента IgG (макрофаги, нейтрофилы, естественные киллеры). Действительно, у больных СПИДом имеются признаки активации комплемента, но это вовсе не означает ее избирательной направленности против зараженных вирусом клеток: динамика ВИЧ-антител не совпадает с сокращением Т-хелперной субпопуляции. Что касается антителозависимой клеточной цитотоксичности, то для ее реализации необходимо столько условий, что многие сомневаются в действенности данного механизма in vivo.
Обсуждается вопрос о ВИЧ-индуцированном апоптозе Т-лимфоцитов. Полагают, что таким способом могут погибать незараженные CD4 клетки, попадающие в сферу влияния ВИЧ-продуктов. Среди факторов, возбуждающих апоптоз, называются сшивка CD4-gp120, блокада клеточного цикла регуляторными ВИЧ-пептидами, сигналы в системе Fas(CD95)—FasL. В инвитровых культурах Т-клетки, обработанные ВИЧ или его продуктами, неадекватно реагируют на антигены и цитокины, подвергаясь апоптозу.
Гистологический анализ обнаруживает глубокое извращение цитоархитектоники лимфатических узлов больных СПИДом, прежде всего за счет разрушения сети дендритных клеток, необходимых для стабилизации межклеточных контактов и представления антигенов Т-лимфоцитам. Не исключено, что это одно из последствий Т-хелперного дефицита, хотя среди изобилия ВИЧ-мутантов встречаются клоны, способные реплицироваться в дендритных клетках и вызывать их повреждение. Это содействует гибели Т-лимфоцитов, выступая в роли кофакторов иммунодефицита. Не следует забывать и о том, что ВИЧ заражает плюрипотентные стволовые клетки костного мозга, тимоциты и стромальные клетки тимуса. Это предрасполагает к ослаблению гемопоэза и тимопоэза, углубляя дефицит иммунокомпетентных клеток и панцитопению, характерную для терминальной фазы заболевания.
Уже в первых публикациях высказывалось мнение о том, что иммунная система при ВИЧ-инфекции исходно страдает не столько из-за дефицита иммунокомпетентных клеток, сколько от нарушения межклеточных контактов. Эта парадоксальная позиция имеет тем не менее серьезные основания. К примеру, на инфицированных лимфоцитах снижается число CD4-рецепторов, необходимых для HLA-зависимых контактов с антигенпредставляющими клетками. Кроме регуляторных ВИЧ-пептидов этому способствует экранирование CD4 свободным белком gp120, который отщепляется от зараженных клеток. Межклеточная кооперация ослабевает и из-за сокращения молекул HLA-2 на поверхности макрофагов и дендритных клеток, что отрицательно сказывается на представлении антигенов Т-хелперам. Вырисовывается и вероятность функциональной блокады CD8 Т-лимфоцитов. Есть данные о том, что этот дефект возникает в начале инфекции, прогрессируя вместе со снижением числа циркулирующих CD4 Т-лимфоцитов и клиникой заболевания.
Много внимания уделяется перекосам в системе цитокинов. В опытах с моноцитами и Т-клетками некоторые из цитокинов (ИЛ-1b, ИЛ-6, гранулоцитарно-макрофагальный колониестимулирующий фактор, a-фактор некроза опухолей) усиливают ВИЧ-репликацию. Их главным источником служат CD4 Т-лимфоциты и макрофаги, которые активируются на фоне ВИЧ- индуцированного цитокинового дисбаланса.
Теперь, пожалуй, следует рассказать о самом щекотливом парадоксе ВИЧ-иммунодефицита. Классическая трактовка СПИДа как финала опережающего обвала Т-хелперов не объясняет дихотомии (расщепления) иммунодефицитного синдрома, а именно избирательного поражения клеточного иммунитета при относительном благополучии гуморального звена. Это трудно понять, если учесть, что индукция гуморального ответа тоже зависит от CD4 Т-клеток и должна была бы страдать не меньше, чем клеточный иммунитет. Выйти из этого противоречия можно, лишь признав неоднородность Т-хелперов с их разделением на варианты, неодинаковые по чувствительности к ВИЧ и по активности в реакциях клеточного и гуморального иммунитета. Подобная мысль была высказана вскоре после обнаружения первых случаев СПИДа. В иммунных реакциях задействовано более 40 цитокинов, часть из которых выделяется CD4 Т-хелперами. Некоторые из них (цитокины 1-го типа) усиливают реакции клеточного иммунитета, другие (цитокины 2-го типа) — образование антител. Т-хелперы, индуцирующие клеточный иммунитет, получили акроним Th1 (от англ. helper); продуценты В-цитокинов называются Th2. Все Th-варианты возникают из Th-предшественников (Th0) под влиянием дифференцировочных цитокинов, исходящих главным образом из самих иммунокомпетентных клеток.
Признание поляризации хелперных Т-лимфоцитов (то же самое справедливо и для CD8 Т-клеток) — одно из крупных откровений последних лет, которое оказало существенное влияние на представления о патогенезе инфекционных и иммунопатологических процессов. Баланс Th1-Th2 динамичен, сдвигаясь в ту или иную сторону при различных нарушениях гомеостаза, в том числе инфекционной природы. Выходя за определенные рамки, дихотомия Т-хелперов может извратить эффекторные функции иммунитета, направляя их в русло патологии. Отсюда не случайно, что ВИЧ-инфекция с характерной для нее поляризацией иммунодефицита оказалась в центре дискуссий относительно патогенетической значимости Th1-Th2 перестроек.
Есть мнение, что ВИЧ индуцирует перекос в пользу Th2-зависимых реакций, одновременно подавляя Th1-зависимую наработку цитотоксических (анти-ВИЧ) Т-лимфоцитов. Это снижает контроль над вирусом, создавая предпосылки для развития СПИДа. Более того, опираясь на медиаторный дисбаланс, некоторые из цитокинов типа 2 способны усиливать репликацию ВИЧ. Преобладание Th2-функций рассматривается как одна из причин лимфаденопатии и гипергаммаглобулинемии — типичных спутников предспидового периода ВИЧ-инфекции (см. ниже). И то и другое может быть результатом избыточной стимуляции В-лимфоцитов в разбалансированной системе цитокинов.
Вместе с тем анализ Th1-Th2 перекосов не дал однозначных результатов. Нет уверенности и в их патогенетическом лидерстве. Не исключено, что это следствие истощения лимфоидной ткани, а не пусковой фактор грядущего иммунодефицита. Однако в любом случае раскрытие причин, сдвигающих равновесие между субпопуляциями CD4 Т-лимфоцитов, намечает перспективы по сдерживанию ВИЧ-патологии на основе исправления цитокиновых перекосов.
Патология
В клинических классификациях детально анализируются стадии и подстадии ВИЧ-инфекции. Богатая клиническая симптоматика вначале отражает конфликт между полноценной иммунной системой и вирусом (острая ВИЧ-инфекция), затем — длительную фазу относительной компенсации (хроническая ВИЧ-инфекция) и, наконец, глубокий иммунодефицит (СПИД). Подготовка к СПИДу занимает около 10 лет, но может быть короче или длиннее. Изредка встречаются случаи с устойчивым уровнем CD4 Т-лимфоцитов, низкой репликативной активностью вируса и полной клинической латенцией. Они остаются интригующей загадкой, составляя менее 5% от общего числа ВИЧ-инфицированных.
Можно говорить о двух основных периодах ВИЧ-инфекции — до и после кризиса иммунной системы. Этому соответствует относительное благополучие (предспидовый период) и катастрофическое ухудшение состояния больного (СПИД). Начало кризисной ситуации знаменует падение CD4 Т-лимфоцитов в крови до 200—300 клеток/мкл. Параллельно ослабевает, а затем исчезает способность к замедленным (Т-зависимым) аллергическим реакциям. Именно с этого момента возникают серьезные проблемы с оппортунистическими инфекциями, спектр и агрессивность которых нарастают к финалу. Одновременно повышается склонность к злокачественным опухолям, прежде всего к саркоме Капоши и В-лимфомам. Лимфоцитарное истощение и разрушение цитоархитектоники лимфоидной ткани ведет к инволюции лимфатических узлов (атрофия и склероз).
В предспидовом периоде клинические проявления во многом связаны с гиперстимуляцией иммунокомпетентных клеток и перекосами в цитокиновом профиле. Острый ВИЧ-синдром обычно начинается спустя три-четыре недели после заражения. По времени он совпадает с первым пиком вирусемии и самокупируется через 2—4 нед на фоне сероконверсии. Спектр возможных симптомов необычайно широк, затрагивая практически все системы организма (табл. 1). Чаще всего возникает мононуклеозоподобный синдром с лихорадкой, ангиной, увеличением лимфоузлов, миалгией и сыпью. Иногда доминируют признаки атипичной пневмонии, гастроэнтерита, серозного менингита. Нередко заражение протекает бессимптомно или, по крайней мере, не обращает на себя серьезного внимания (первичная латенция).
Таблица 1
Возбудители СПИД-индикаторных инфекций
| Простейшие | Грибы | Бактерии | Вирусы |
| Toxoplasma gondii | Pneumocystis carinii | Salmonella spp. | Herpes simplex virus |
| Isospora belli | Candida albicans | Mycobacterium avium complex | Cytomegalovirus hominis |
| Cryptosporidium spp. | Cryptococcus neoformans | Mycobacterium tuberculosis | |
| Histoplasma capsulatum | |||
| Coccidioides immitis |
После острой стадии наступает внешнее затишье. Оно символизирует не более чем временный контроль над репликацией вируса, требующий серьезного напряжения защитных ресурсов. Это нередко проявляется в виде длительного (месяцы, годы) увеличения лимфоузлов — персистирующая генерализованная лимфаденопатия. Она отражает гиперплазию лимфоидных фолликулов за счет гиперстимуляции В-лимфоцитов непрерывно реплицирующимся вирусом и В-лимфотропными цитокинами. Возможна и активация вируса Эпстайна—Барр, который персистирует в В-лимфоцитах многих людей и обладает мощным В-стимулирующим потенциалом. Этому способствует дефицит контролирующих сигналов со стороны Т-лимфоцитов. Подвергаясь неспецифической стимуляции, В-лимфоциты работают вхолостую, продуцируя бесполезные иммуноглобулины. Сама по себе лимфаденопатия не коррелирует с прогнозом и не приближает катастрофы. Примерно у трети больных она протекает бессимптомно и обнаруживается при случайном осмотре. В остальных случаях болезнь заявляет о себе периодическими подъемами температуры, приступами слабости, потливости и пр.
Начало СПИДа знаменует новый виток ВИЧ-инфекции. Болезнь складывается из суммы вторичных инфекций и злокачественных опухолей, манифестирующих ВИЧ-индуцированную гибель иммунной системы. Вклад самого вируса на этом фоне выглядит скромно. Среди возбудителей оппортунистических инфекций преобладают внутриклеточные паразиты, устойчивость к которым зависит от клеточного иммунитета. Они поступают в организм извне или оживают при активации эндогенной инфекции (табл. 1). К группе так называемых СПИД-индикаторных, или СПИД-ассоциированных инфекций относится 13 нозологических единиц, хотя этот список может быть расширен (табл. 2).
Таблица 2 СПИД-индикаторные болезни
|
Болезни |
| 1. Множественные или рекуррентные бактериальные инфекции у детей младше 13 лет: септицемия, пневмония, менингит, инфекция суставов и костной ткани или абсцессы внутренних органов, связанные с палочкой инфлюэнцы, стрептококками или другими пиогенными бактериями |
| 2. Кандидоз пищевода, трахеи, бронхов или легких |
| 3. Диссеминированный кокцидиодомикоз |
| 4. Экстрапульмонарный криптококкоз |
| 5. Хронический криптоспоридиоз с диареей продолжительностью более 1 мес |
| 6. Цитомегаловирусная инфекция |
| 7. Герпетическая инфекция слизистых оболочек и кожи, персистирующая более 1 мес |
| 8. ВИЧ-энцефалопатия |
| 9. Диссеминированный гистоплазмоз |
| 10. Изоспориоз с диареей продолжительностью более 1 мес |
| 11. Саркома Капоши |
| 12. Первичная лимфома головного мозга |
| 13. Неходжкинская лимфома В-клеточного или неизвестного фенотипа (включая лимфому Беркитта) |
| 14. Лимфоидная интерстициальная пневмония у детей младше 13 лет |
| 15. Диссеминированная микобактериальная инфекция (не М. tuberculosis) |
| 16. Экстрапульмонарный туберкулез |
| 17. Пневмоцистная инфекция |
| 18. Прогрессирующая многоочаговая лейкоэнцефалопатия |
| 19. Рецидивирующая сальмонеллезная инфекция |
| 20. Токсоплазмоз головного мозга |
Склонность к злокачественным опухолям (причем таким редким, как саркома Капоши и В-лимфомы) тоже логично вписывается в иммунодефицитный синдром. Сам по себе ВИЧ лишен онкогенности, но, ослабляя иммунологический надзор, выводит из-под контроля другие, опасные в этом отношении вирусы или повышает злокачественный потенциал трансформированных цитокинами клеток. Гиперстимуляция цитокинами рассматривается как один из вероятных механизмов усиления ангиногенеза, необходимого для развития саркомы Капоши и В-лимфом. Это может повышать чувствительность к другим канцерогенам, в частности к вирусам. Много дебатов ведется вокруг новых герпесвирусов, которые впервые были изолированы из В-лимфом и саркомы Капоши больных СПИДом. Герпесвирус-8 признан основным претендентом на роль возбудителя саркомы Капоши (см. «Герпесвирусы»).
После первых признаков СПИДа продолжительность жизни больных не превышает 5 лет.
Неврологические проявления СПИДа. ВИЧ проникает в нервную ткань вскоре после заражения. Головной и спинной мозг могут страдать от лимфоидных опухолей или инфекций, чаще всего от Cryptococcus neoformans (менингит) и Toxoplasma gondii (внутримозговые абсцессы). Но у ряда больных неврологические симптомы развиваются независимо от вторичных осложнений, свидетельствуя о прямой нейротропности самого вируса. Это не должно удивлять, так как для «родственников» ВИЧ (медленных вирусов животных) поражение ЦНС — одно из центральных проявлений болезни. Главным оплотом для ВИЧ служат клетки микроглии — миниатюрные макрофаги ЦНС, которые, как и все тканевые макрофаги, возникают из моноцитов крови. Скорее всего именно моноциты доставляют вирус в ЦНС, активно преодолевая гематоэнцефалический барьер.
Преходящие неврологические симптомы (менингит, энцефалит, миелит, полиневриты) иногда возникают в остром периоде ВИЧ-инфекции, и, возможно, на этом этапе закладываются вирусологические основы для необратимого поражения ЦНС. Начинаясь с малых признаков (забывчивость, рассеянность, заторможенность, нарушение почерка, психическая депрессия), СПИД-энцефалопатия ведет к слабоумию и двигательно-вегетативным расстройствам, приковывающим к постели. Это так называемый нейро/психо-СПИД, который столь же неумолимо, как инфекто-СПИД и онко-СПИД, ведет к смерти. В большинстве случаев он сочетается с другими признаками, включаясь в симптомокомплекс полного СПИДа 1.
Механизм поражения ЦНС не ясен. Известно лишь, что ВИЧ ведет к реактивной пролиферации глиальных клеток и кортикальной атрофии. Такие признаки обнаруживаются у 75% больных, погибших от СПИДа. Для этого вирус не обязан размножаться в нейронах. Достаточно gp120-зависимой блокады рецепторов для нейропептидов, которая разобщает клеточное сообщество, лишая его тонизирующих сигналов.
Интестинальные проявления СПИДа. Хроническая диарея — один из обычных симптомов СПИДа. Часто это обусловлено вторичной инфекцией протозойной (криптоспоридии, изоспоры, лямблии), бактериальной (сальмонеллы, атипичные микробактерии, кампилобактеры, шигеллы) и вирусной (цитомегаловирус) природы. Но нередко установить этиологию не удается. Возможно, сам ВИЧ служит причиной энтеропатии, поражая эпителий кишечника. Длительная диарея ведет к нарушению всасывания и истощению организма. Больной резко теряет в весе, оправдывая слэнговый синоним СПИДа — болезнь похудания (англ. slim disease). Под таким названием СПИД известен в Африке, где ВИЧ-кахексия встречается особенно часто и протекает наиболее тяжело. Причины этого не ясны. Возможно, болезнь вызывает не сам ВИЧ, а неизвестный микроорганизм, распространенный исключительно в Африке.
Лабораторная диагностика
Диагноз основан на выявлении вирусспецифических антител. Для уточнения можно исследовать сыворотку на присутствие ВИЧ-антигенов (р24) и РНК (рис. 6). Определение антител проводят при помощи высокочувствительных реакций иммуноферментного анализа (ИФА). В качестве ВИЧ-антигенов применяют производные env- и gag-комплекса. Сегодня уже не пользуются материалом из вирусных культур, а предпочитают рекомбинантные или синтетические пептиды. Обнаружение антител говорит о заражении вирусом и эпидемической опасности его носителя. Впрочем, всегда остается небольшая надежда на гипердиагностику, причиной которой могут быть случайные антитела, перекрестно реагирующие с ВИЧ-антигенами.

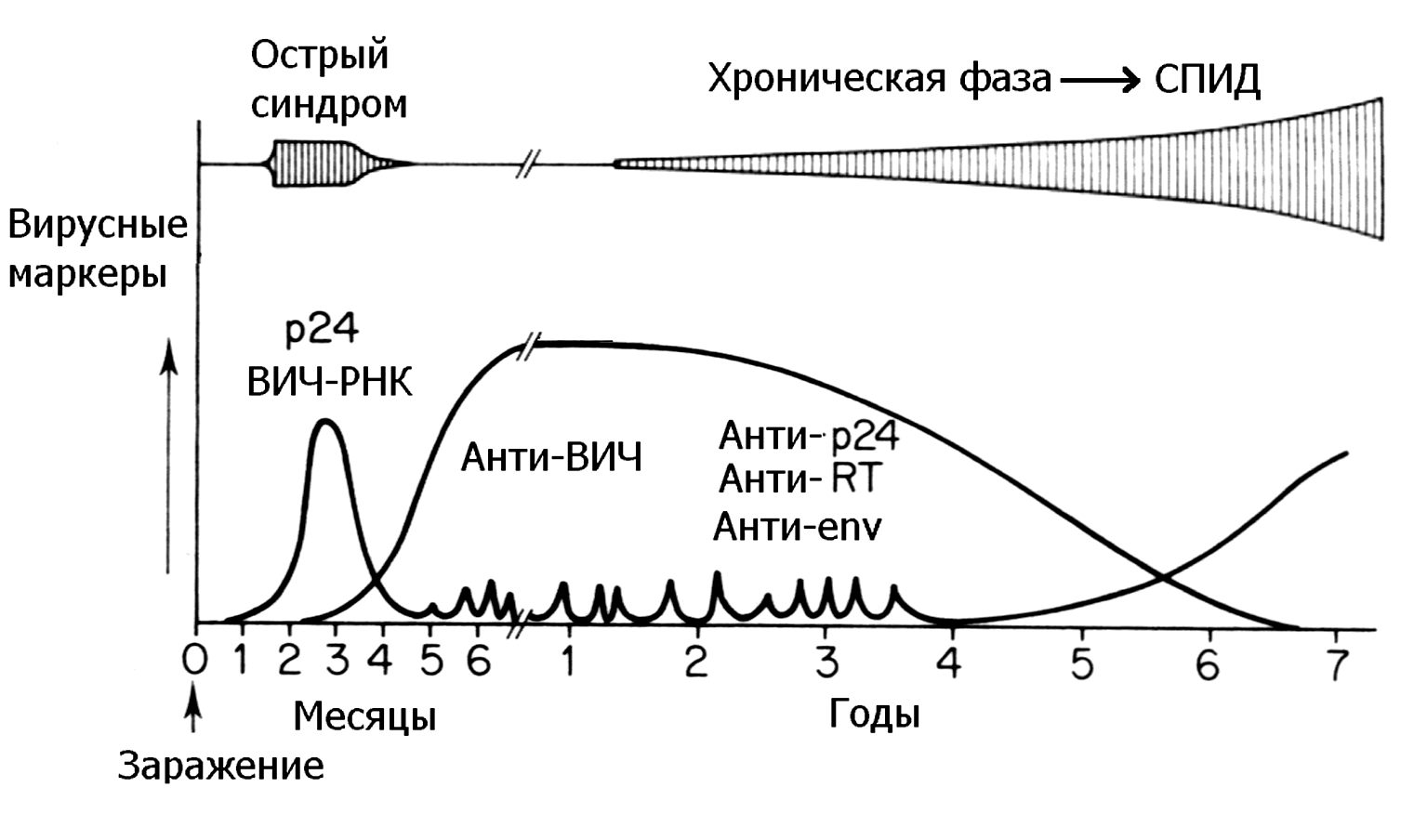
Ложную тревогу снимают при помощи иммуноблоттинга. В отличие от обычного ИФА, иммуноблоттинг не только выявляет антитела по принципу да-нет, но позволяет дифференцировать реакции с отдельными антигенами, которые предварительно рассортировывают на носителе (обычно нитроцеллюлозе) путем электрофореза. После обработки полоски носителя исследуемой сывороткой проводят обычный ИФА, т.е. проявляют связавшиеся с антигенами антитела при помощи меченых антииммуноглобулиновых антител.
Положительные реакции выглядят в виде ИФА-пятен (англ. blot — пятно), каждое из которых соответствует дискретной паре антиген-антитело. Единственное пятно позволяет усомниться в истинности результата. Картину проясняет повторный анализ, сделанный через несколько месяцев. Усложнение спектра антител говорит о ВИЧ-инфекции, отсутствие динамики доказывает ложную позитивность первичного ИФА (рис. 7).
Дополнительно к иммуноблоттингу (или вместо него) для решения спорных вопросов пользуются прямыми ВИЧ-маркерами — присутствием в крови главного нуклеокапсидного белка (p24) и ВИЧ-генома (см. рис. 6). р24 определяют путем ИФА, ВИЧ-РНК — при помощи полимеразной цепной реакции.

Встречается и ложная серонегативность. Чаще всего это наблюдается в раннем периоде ВИЧ-инфекции в связи с отсроченной сероконверсией. Антитела обычно появляются в течение первых трех месяцев и могут отсутствовать, несмотря на вирусемию. Это так называемое диагностическое окно, продолжительность которого иногда затягивается на полгода и более (в редких случаях до года). Антитела нередко исчезают в терминальной фазе СПИДа, когда разрушение иммунной системы захватывает и ее гуморальное звено. В подобных случаях можно положиться на маркеры ВИЧ-репликации — р24 и РНК. Впрочем, к этому прибегают редко, так как можно либо подождать до появления антител, либо серопозитивность уже зафиксирована в анамнезе.
Антигенные и генетические различия между ВИЧ-1 и ВИЧ-2 требуют специальных для каждого из них тест-систем. Они могут быть объединены в общие диагностикумы, но суть от этого не меняется.
Принципы антивирусной терапии и проблемы вакцинации
Блокировать ВИЧ-репликацию научились вскоре после открытия вируса. Это удалось сделать при помощи аномальных нуклеозидов, подавляющих активность вирусспецифического фермента — обратной транскриптазы. Все ингибиторы подобного типа относятся к группе так называемых терминаторных субстратов, поскольку их эффект связан с обрывом (терминацией) синтеза ДНК на вирусной РНК-матрице. Наиболее известен азидотимидин (АЗТ; коммерческие названия — зидовудин, ретровир, тимазид), который до недавних пор был единственным средством, продлевающим жизнь больных СПИДом. Позже появились его клинические аналоги — ставудин, ламивудин, видекс, залцитабин и др.
Первый ажиотаж вокруг АЗТ быстро прошел. Виной этому оказалась высокая мутабельность вируса. На протяжении 6 мес в ВИЧ-популяции возникают разистентные клоны, которые захватывают лидерство в инфекционном процессе, сводя на нет эффективность терапии. Замена АЗТ на родственные препараты не достигает цели, так как все они блокируют один и тот же участок обратной транскриптазы, а потому попадают под общий механизм мутационной устойчивости.
В 1990 г. стали известны соединения ненуклеозидной природы, эффективно подавляющие обратную транскрипцию ВИЧ-1. Принципиально, что они взаимодействуют с участком молекулы, который, хотя и расположен по соседству с сайтом связывания для ингибиторов группы АЗТ, но мутирует независимо от него. Поэтому резистентность к АЗТ не распространяется на ненуклеозидные ингибиторы обратной транскриптазы (ННИОТ). К сожалению, устойчивость к ННИОТ (например, к невирапину, ловириду, эфавирену и др.) формируется едва ли не быстрее, чем к нуклеозидным ингибиторам. Преимуществом является то, что привыкание в этом случае не носит универсального характера, т.е. резистентность к одному из ННИОТ не всегда распространяется на другие. В отличие от нуклеозидных ингибиторов ННИОТ действуют только на ВИЧ-1 и неэффективны против ВИЧ-2. Следуя логике, можно думать, что вирусу труднее выжить в среде с двумя разнотипными ингибиторами подобно тому, как развитие лекарственной резистентности у бактерий замедляется при сочетанном применении разных антибиотиков. Эта идея стимулировала разработку комплексной анти-ВИЧ-терапии, которая пришла на смену монотерапии нуклеозидными ингибиторами.
Дальнейшее развитие сочетанной анти-ВИЧ-терапии связано с торможением вирусспецифической протеазы. Как уже говорилось, этот фермент необходим для репликации вируса, превращая полипротеины в зрелые вирионные белки. Синтезировано множество соединений, блокирующих протеолитическую активность ВИЧ. Почти все они действуют по принципу пептидомиметиков, имитируя пептидную связь, атакуемую ВИЧ-протеазой. Протеазные ингибиторы (саквинавир, ритонавир, индинавир, нельфинавир, ампренавир, лопинавир и др.) вызывают быстрое и значительное снижение вирусной нагрузки. Недостаток прежний: продолжительность эффекта ограничена привыканием вируса. Уже через 6—12 мес после начала лечения в организме больных появляются устойчивые ВИЧ-варианты, причем резистентность, индуцированная одним препаратом, распространяется и на другие ингибиторы этого типа.
Таким образом, сегодня известны три группы клинически значимых ингибиторов ВИЧ-репликации. Они используют разные мишени и поэтому не вызывают перекрестной устойчивости вируса друг к другу. Кроме того, внутри одной из групп (ННИОТ) перекрестное привыкание возникает не во всех сочетаниях. Современные схемы (они получили название активного, даже агрессивного лечения) предусматривают тройную терапию на основе комбинации двух (нуклеозидного и ненуклеозидного) ингибиторов обратной транскриптазы и одного из антипротеазных препаратов. Применение такого коктейля ведет к быстрому падению вирусемии вплоть до уровня, не уловимого полимеразной цепной реакцией. Предлагаются и более громоздкие варианты — четырех и даже пяти препаратов. Усложнение идет за счет ННИОТ, не вызывающих перекрестного привыкания вируса.
Лечение рекомендуется начинать как можно раньше и продолжать возможно дольше. Для слежения за эффективностью терапии используют маркеры вирусной репликации, прежде всего количественные показатели РНК-вирусемии. Повышение титра ВИЧ-РНК говорит о необходимости возобновления или интенсификации лечения.
К сожалению, надежда на то, что многокомпонентная терапия обеспечит полную ВИЧ-стерилизацию организма, не оправдалась. ВИЧ сохраняет свой непредсказуемый мутационный потенциал тем более, что избирательность ВИЧ-ингибиторов не абсолютна, и при напряженной терапии сказывается их токсичность. Кроме того, интенсификация терапии делает ее дорогостоящей, о чем задумываются даже в экономически развитых странах.
И все-таки прогресс налицо. Он блистательно отражает связь между теорией и практикой, между тонкостями молекулярной биологии и клинической медициной. Кстати, задействованы еще не все вирусспецифические мишени, что вселяет надежду на очередные всплески ВИЧ-химиотерапии.
Идея создания ВИЧ-вакцины возникла тотчас после открытия вируса и поначалу не сулила серьезных проблем. Возбудитель известен, культивируется, его антигены проанализированы вплоть до протективных эпитопов, гены клонированы и готовы к наработке любых количеств высокоочищенного вирусного материала. Словом, казалось, что, опираясь на современную технологию, создать надежную вакцину будет несложно. Но годы напряженного труда не привели к желаемому результату. Причин несколько. Прежде всего следует помнить о высокой антигенной изменчивости суперкапсидных гликопротеинов ВИЧ, прежде всего gp120. Опыты с нейтрализацией вируса в культурах клеток и на ВИЧ-инфицированных шимпанзе показывают, что протективный эффект против лабораторных штаммов не гарантирует защиты от клинических (диких, или полевых) изолятов. Об этом же говорят и единичные (но принципиальные!) наблюдения о возможности заражения ВИЧ несмотря на вакцинацию. Необходимы широкомасштабные исследования, чтобы сопоставить численность таких случаев с контрольной группой, но организовать такие испытания непросто.
Проверка эффективности ВИЧ-вакцин сопряжена с большими трудностями. Экспериментальные модели, дублирующие инфекцию человека (СПИД обезьян и кошек), приносят пользу, но имеют очевидные ограничения. Шимпанзе восприимчивы к ВИЧ, но клинически инфекция у них не проявляется. Исследования на клеточных культурах позволяют изучать базисные закономерности инфекции, но далеки от клинических реалий. Испытания на людях сопряжены с серьезными этическими проблемами.
Не ясен вопрос о возможности унификации вакцин при разных способах инфицирования. К примеру, вакцины на основе оболочечных антигенов предупреждают развитие инфекции при внутривенном заражении (опыты на шимпанзе), но длительность эффекта и его надежность при мукозальной инвазии (половой путь) остаются неясными. Препятствием для антител может быть вирус (провирус), проникающий в организм внутри клеток. Дополнительные проблемы связаны с вирусом, транспортируемым в лимфоидную ткань дендритными клетками.
Большинство попыток изготовить вакцину сконцентрировано вокруг суперкапсидных антигенов, главным образом gp120. Испытан или испытывается весь арсенал технических средств современной вакцинологии: 1) рекомбинантные белки, получаемые в различных генно-инженерных системах, 2) синтетические пептиды, аналогичные иммунодоминантным эпитопам, 3) векторные вакцины, несущие ВИЧ-гены (например, рекомбинантные поксвирусы, содержащие env-, gag- и pol-гены), 4) ДНК-вакцины, сконструированные на основе бактериальных плазмид, дополненных ВИЧ-генами. Появилось больше смелости в испытании различных вакцинных препаратов, схем их сочетанного и последовательного применения.
В последние годы наметился отход от антител как единственного критерия иммуногенности вакцин. Появился интерес к Т-вакцинам, которые стимулируют клеточный иммунитет, реализуемый цитотоксическими Т-лимфоцитами. Мы уже говорили о том, что при ВИЧ-инфекции они появляются раньше вируснейтрализующих антител, сдерживая репликацию вируса. Более того, есть данные, что цитотоксические Т-лимфоциты способны полностью уничтожить ВИЧ-содержащие клетки, искоренив инфекцию в самом начале. Об этом говорят наблюдения за лицами из групп риска, которые несмотря на повторные контакты с ВИЧ остались незараженными. У таких людей серонегативность сочетается с наличием анти-ВИЧ CD8 Т-лимфоцитов.
Кроме поверхностных гликопротеинов, Т-активностью обладают внутренние белки вириона и даже регуляторные пептиды. Т-эпитопы более консервативны, чем структуры, возбуждающие синтез антител, а, значит, и вакцины, полученные на их основе, будут более универсальны, т.е. менее зависимы от изменчивости вируса. Не исключено, что для воспроизведения полноценной защиты потребуется сочетание разных ВИЧ-компонентов. Казалось бы, не надо мудрствовать: лучше всего должна работать вакцина из цельных вирионов — убитых или аттенуированных. И то и другое проверялось, но тоже с неоднозначным результатом. К тому же страх перед остаточной инфекционностью ВИЧ настолько велик, что вирионные вакцины вызывают опасение.
Наиболее азартные замыслы нацелены на создание не только превентивной, но и лечебной вакцины. Она должна индуцировать стерилизующий иммунитет или хотя бы сдерживать ВИЧ-репликацию, как это происходит в начале инфекционного процесса. Иными словами, ставится задача искусственного продления фазы относительного иммунитета, быстро слабеющего на фоне ВИЧ-агрессии. По этому поводу высказано немало сомнений, прежде всего относительно возможности спасения больных с далеко зашедшей патологией, когда повреждение иммунной системы может быть необратимым, а реактивность к антигенам отсутствует. Если пессимистические прогнозы все же не сбудутся, то с технической точки зрения терапевтические вакцины не будут отличаться от профилактических препаратов. Нужно лишь смелее опираться на инактивированные и аттенуированные вирусы, так как остаточная инфекционность в данном случае не ухудшит ситуацию.
Медицинская микология
Жизнь есть везде, но везде есть и враги жизни.
В.И. Вернадский
- Классификационные и структурные сложности.
- Диалектика диформных грибов.
- Совершенные грибы и способы полового размножения.
- Несовершенные грибы, или дейтеромицеты.
- Сапрофитизм как основа экологии грибов.
- Фитопатогенные грибы и оппортунистические микозы.
- Факторы антифунгальной защиты.
- Микоаллергозы и микотоксикозы.
Открытие болезнетворности грибов на несколько лет опередило пионерскиерабо ты Пастера и Коха с патогенными бактериями. В 1839 г. Шенлейн и Граби установили фунгальную (грибковую) природу парши (Trichophyton schoenleini), и в том же году Ландженбек (Langenbeck) обнаружил дрожжеподобные микроорганизмы (Candida albicans) при молочнице. Первый возбудитель системных микозов (кокцидиоидомикоз — Coccidioides immitis) был открыт в 1892 г. Посадас (Posadas) в Аргентине.
Несмотря на ранние работы, медицинская микология долго оставалась в тени бактериологии и вирусологии, хотя грибковые болезни относятся к числу наиболее распространенных инфекций человека. Недостаток внимания можно объяснить относительной доброкачественностью микозов и редкостью (часто географической эндемичностью) тяжелых заболеваний.
Ситуация изменилась в последние десятилетия. Началом стало широкое использование антибиотиков, породившее проблему кандидомикоза (кандидоза), ранее известного, по сути, лишь в виде молочницы у новорожденных. С внедрением в клиническую практику лучевой терапии, стероидных гормонов, иммунодепрессантов, цитотоксических агентов, парентерального питания и протезирования возникла проблема оппортунистических микозов, которые заявили серьезные претензии на «почетное место» среди госпитальных инфекций. Положение заметно обострилось в начале 1980-х гг. в связи с эпидемией СПИДа. Стало очевидным, что системные (глубокие) микозы принадлежат к числу наиболее тяжелых осложнений у иммунокомпрометированных больных.
Грибы относятся к эукариотам, т.е. по строению клеток сходны с растениями и животными. Это означает, что их клетки структурированы (компартментализованы) благодаря системе внутриклеточных мембран, которые формируют морфологически оформленное ядро, разветвленный эндоплазматический ретикулум, митохондрии и другие органеллы. Ядро содержит набор хромосом, реплицирующихся путем митоза. Подобно всем эукариотам для плазматической мембраны грибов характерно высокое содержание стеролов, главным образом эргостерола. Кроме того, грибы способны к половому размножению. Все грибы — аэробы, и лишь немногие способны выживать за счет брожения.
Грибы устроены гораздо примитивнее, чем классические (высшие) эукариоты. Это проявляется прежде всего в низкой специализации клеток, из которых они состоят. Даже у многоклеточных грибов, например плесеней, каждая отдельная клетка способна дать начало целому организму. В отличие от высших эукариот большинство грибов гаплоидны; из «медицинских» грибов диплоидны только Candida. В связи с этим наряду с простейшими и водорослями грибы называют протистами, т.е. низшими эукариотами.
В старых классификациях грибы относили к бесхлорофильным растениям. Это сходство оказалось формальным, так как по ряду базисных признаков грибы радикально отличаются от растений. Они являются хемотрофами, извлекая энергию из химических связей и поэтому великолепно растут в темноте; растения — фототрофы, преобразующие энергию солнечного света в макроэрги (АТФ) путем фотосинтеза. Грибы — гетеротрофы, т.е. их метаболизм базируется на утилизации органических соединений, обычно мертвого органического материала (большинство грибов — сапрофиты); растения — автотрофы, живущие за счет неорганических форм углерода (СО2) и других органогенов.
Это послужило основанием для выделения грибов в самостоятельное царство эукариот — Mycota (греч. mykes — гриб), которое насчитывает около 250 000 видов. По другим данным, эта цифра значительно ниже. Например, в 9-м издании известного руководства «Topley & Wilson’s Microbiology and Microbial Infections» сообщается о 72000 видов. Кстати, из всех предыдущих изданий (руководство выходит, начиная с 1929 г.) вопросы микологии нашли отражение только в последнем (1999 г.), где им посвящен специальный объемистый том.
Для человека болезнетворны примерно 150 грибов. Они вызывают заболевания, называемые микозами. Некоторые грибы продуцируют сильные токсины, опасные для человека и животных. Отравления микотоксинами именуются микотоксикозами. Наконец, продукты грибов могут сенсибилизировать человека, предрасполагая к развитию аллергических заболеваний (микоаллергозы).
Снаружи клетки грибов окружены плотной многослойной оболочкой — клеточной стенкой. Ее основу образуют микрофибриллы, построенные из полисахаридов, главными из которых являются глюканы, маннаны и хитин. Соотношение отдельных полисахаридов варьирует не только у разных видов грибов, но и зависит от фазы онтогенеза (например, различается в дрожжевых и гифальных элементах диморфных грибов). Полисахариды составляют 80—90% клеточной стенки, остальное приходится на белки, липиды и полифосфаты, цементирующие полисахаридный матрикс. Целлюлоза встречается редко, что отличает грибы от растений.
Классификация и размножение
Согласно одной из классификаций, грибы принято делить на три группы: 1) дрожжи, 2) плесени и 3) шляпочные грибы. Возбудители микозов относятся к двум первым группам, и именно о них пойдет речь ниже.
Деление на дрожжи (Blastomycetes) и плесени (Hyphomycetes) основано на двух простых, но принципиальных критериях:
1. Культуральные особенности. Колонии дрожжей похожи на бактериальные — гладкие, пастообразные, вырастают через 24—48 ч. Рост плесеней известен всем из бытовых наблюдений. Это пушистые, часто пигментированные колонии, созревающие медленнее дрожжей. Споры, символизирующие зрелость плесневых колоний, обычно появляются через 4—7 дней (рис. 1).


Рис. 1. Колонии грибов: а — дрожжи (C. albicans), б — плесень (Penicillium chrysogenium)
2. Организация клеток в колониях. Дрожжи — в основном изолированые клетки круглой или овальной формы размером 3—10 мкм. Поэтому о них говорят как об одноклеточных организмах. Колония дрожжей представляет скопление огромного числа клеток разной величины (результат почкования). Все дрожжи в этом отношении одинаковы, и их видовая дифференцировка строится главным образом на особенностях метаболизма. Плесени образуют мицелий, который состоит из беспорядочного переплетения тонких нитей — гиф (греч. huphe — сеть). Отсюда другое название плесеней — нитчатые, или филаментозные грибы. Это структурная основа, тело гриба, или таллус (греч. thallоs — росток, зеленый побег).
Гифы представляют собой трубки, наполненные клеточным содержимым; диаметр колеблется от 0,8 до 15 мкм. Обычно они имеют перегородки — признак высших грибов. Такой мицелий называют септированным, т.е. разделенным на клетки. Плесени, у которых мицелий не септирован, по сути представляют единую гигантскую многоядерную клетку. Их называют ценоцитными (от греч. koenos — единый, общий), или низшими грибами. Впрочем, септированный мицелий функционально не отличается от ценоцитного, так как в межклеточных перегородках имеются перфорации, через которые клетки (обычно содержащие несколько ядер) обмениваются друг с другом своим содержимым.
Плесени (особенно высшие) отвечают понятию многоклеточности. Это справедливо, потому что разные отделы мицелия выполняют неодинаковые функции, т.е. гифальные клетки в известной мере специализированы. Подобно растениям, у мицелия есть корни (ризоиды; греч. rhiza — корень), которые закрепляют плесень на поддерживающем субстрате и «кормят» ее тело. Эта часть называется субстратным, или вегетативным мицелием. Остальная (надсубстратная) часть называется воздушным, или репродуктивным мицелием, так как на нем созревают репродуктивные клетки — споры (конидии) (см. ниже). В целом колония плесени напоминает маленькое растение, хотя это сходство имеет чисто внешний характер (рис. 2).

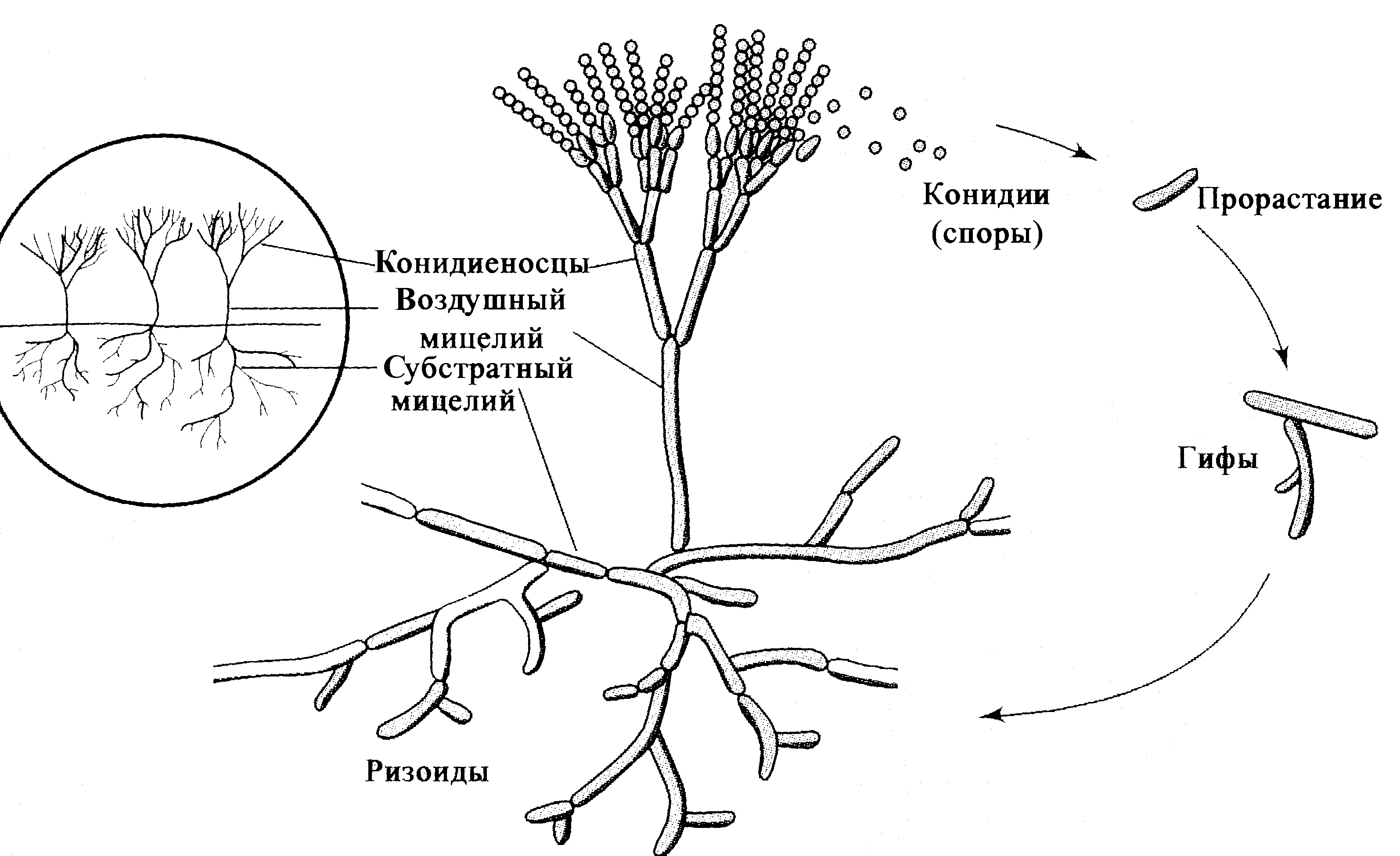
Формально дрожжи и плесени совершенно непохожи. Но есть феномен, который убеждает в противном. Известны грибы, которые в зависимости от условий растут как дрожжи или как плесени. Это явление называется диморфизмом, а такие грибы — диморфными грибами. Диморфизм характерен для возбудителей системных (глубоких) микозов человека — бластомикоза (Blastomyces dermatitidis, Paracoccidioides brasiliens), гистоплазмоза (Histoplasma capsulatum) и кокцидиоидомикоза (Coccidioides immitis). В организме хозяина они образуют дрожжеподобные клетки, а в инвитровых культурах растут в виде мицелиальных форм. Это связано с реакцией на температурный режим, где происходит репродукция грибов (температурозависимый диморфизм). Рост в дрожжевой фазе происходит при 37°С (т.е. при температуре тела человека), комнатная температура инициирует формирование мицелия.
Диморфизм может проявляться и несколько иначе: оба морфологических варианта сосуществуют в зараженном организме. Это характерно для Candida albicans — возбудителя одного из самых распространенных микозов человека. Обычно C. albicans представлены дрожжеподобными клетками, которые могут формировать цепи — псевдогифы. Но одновременно формируются и истинные гифы (трубки без перетяжек, т.е. с общей стенкой) (рис. 3). Подобно гифам плесневых грибов, их образование начинается с ростковых трубок, которые, удлиняясь, превращаются в длинные нити (рис. 4). Одним из факторов, стимулирующих образование ростковых трубок (и далее гиф), служит повышенное содержание СО2 (с такими условиями возбудитель сталкивается в тканях). Феномен легко воспроизводится in vitro и имеет диагностическое значение. Вполне вероятно, что появление ростковых трубок символизирует патогенетически значимую активацию кандид, предрасполагая к инвазии.

Грибы размножаются спорами, которые образуются на одном и том же мицелии неполовым (вегетативным) и половым способами. Грибы, размножающиеся половым путем, считают совершенными (fungi perfecti), те, у которых половой процесс отсутствует, называют несовершенными (fungi imperfecti), или дейтеромицетами (от греч. deuteros — второй).

Это вовсе не означает, что они действительно асексуальны; скорее, половая фаза их жизненного цикла просто не установлена. Если это удается сделать, из несовершенного вид становится совершенным. Это говорит об условности категории «несовершенные грибы», включение в которую часто основано на недостатке информации. Здесь соседствуют совершенно разные грибы только потому, что у них не обнаружен половой процесс. Не случайно о дейтеромицетах (к ним относятся многие грибы, имеющие медицинское значение) говорят как о мусорной корзине или таксономической помойке.
Репродукция неполовыми спорами соответствует анаморфному состоянию; телеморфная фаза — период образования половых спор; сочетание неполового и полового размножения обозначается как голоморфность. Соответственно структуры, обеспечивающие бесполое размножение, называются анаморфами, а плодовые тела, образующиеся в результате полового процесса — телеморфами. Совершенные грибы могут пребывать в любом из трех состояний (ана-, теле- и голоморфном); для несовершенных грибов известна только анаморфность. Несовершенные грибы, перешедшие в ранг совершенных, сохраняют прежнее (анаморфное) название и приобретают новое, которое соответствует телеморфной стадии. Например, старые (анаморфные) родовые названия Trichophyton и Microsporium (возбудители дерматомикозов) в телеморфном варианте заменены на Arthroderma. Впрочем, претензии такого рода пока не увенчались успехом, и прежние названия, полученные из-за неправильных представлений о несовершенстве этих и других видов клинически значимых грибов, по-видимому, долго останутся в употреблении.
Согласно недавним рекомендациям, понятия «совершенные» и «несовершенные» грибы признаны нецелесообразными. Царство грибов предложено подразделить на четыре подцарства (Phyla). Три из них отражают способ образования половых спор (см. ниже): аскомицеты (Ascomycota), базидиомицеты (Basidiomycota) и зигомицеты (Zygomycota). Вместо дейтеромицетов введено понятие «митоспоровые грибы» — термин, отражающий репродукцию исключительно неполовыми спорами, т.е. путем митоза. Будущее покажет жизнеспособность этого (в целом разумного) предложения.
Так или иначе, но половой способ размножения для грибов не обязателен. Безусловно, он стратегически важен как механизм конструирования новых генотипов, но массовость образования вегетативных (неполовых) спор в сочетании с мутациями компенсирует отсутствие генетических рекомбинаций, достигаемых половым общением.
Неполовое размножение. Для неполовых спор грибов принят универсальный термин «конидии» — производное греческого слова konis, пыль. Это отражает легкость, с которой плесневые споры распространяются (распыляются) в окружающей среде.
Дрожжи размножаются почкованием, т.е. путем асимметричного деления клеток (реже наблюдается симметричное деление) (рис. 5). Дочерние клетки (почки, или бластоконидии) быстро отделяются от материнской клетки или, продолжая деление, формируют псевдогифы.

У плесневых грибов дело обстоит сложнее. Главный механизм сводится к созреванию профессиональных конидий, которые в огромном количестве отшнуровываются от специализированных (конидиогенных) гиф репродуктивного мицелия. По сути, это усложненный процесс почкования, напоминающий формирование дрожжевых псевдогиф. У некоторых плесеней (зигомицеты) конидии развиваются внутри сумчатых структур — спорангиев и высвобождаются после их разрыва (спорангиоконидии, или спорангиоспоры). Размеры (3—16 мкм), форма, расположение и цвет конидий, а также строение конидиогенных гиф (конидиофоров, или конидионосцев) являются дифференциальными признаками различных родов и видов плесневых грибов (рис. 6).

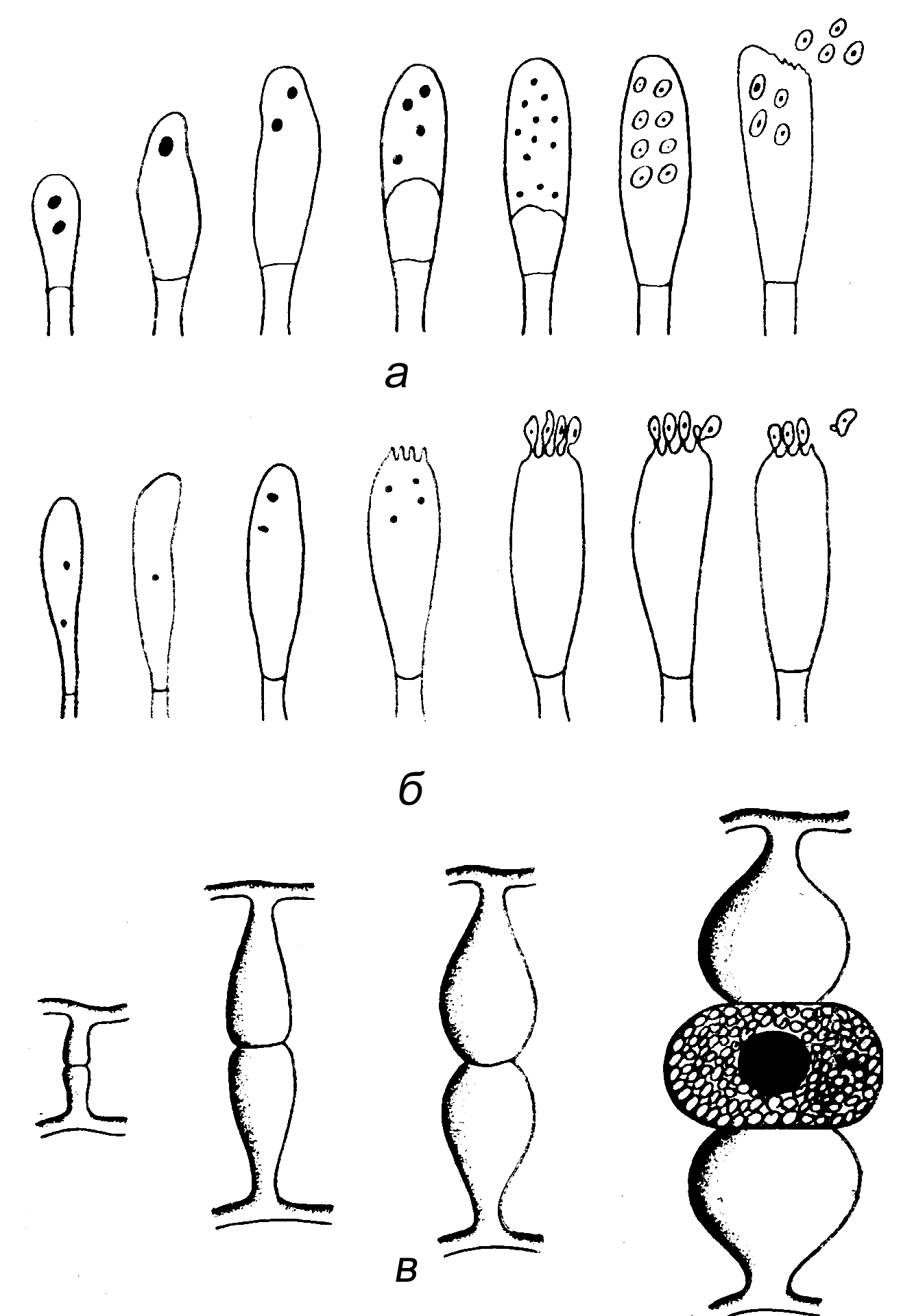
Бесполое размножение плесеней возможно и без образования профессиональных конидий, т.е. спор, созревающих на специализированных структурах. В таких случаях споры образуются почкованием по ходу обычных гиф родительского мицелия (бластоконидии) или путем его фрагментации (артроконидии). Дополнительной разновидностью спор являются хламидоконидии. Они обеспечивают не размножение, а выживание грибов в неблагоприятных условиях. В этом отношении хламидоконидии соответствуют эндоспорам бактерий, хотя и менее устойчивы. Они окружены двойной оболочкой и образуются у большинства грибов.
Некоторые плесени образуют разные типы конидий на одном и том же мицелии. Одни из них (одноклеточные) имеют небольшие размеры и называются микроконидиями, другие содержат несколько клеток — макроконидии. Одиночные микроконидии (бластоконидии) на боковых ответвлениях гиф (они наблюдаются у возбудителей дерматофитозов) называются алейриями (от греч. aleuron — мука).
Половое размножение. Известно три способа образования половых спор у грибов.
Аскоспоры. Их образуют аскомицеты, к которым кроме плесневых грибов относятся многие дрожжи. Проще всего дело обстоит у дрожжей-аскомицетов. Две соседние клетки сливаются через короткие трубкообразные отростки, ядра объединяются, и возникшее диплоидное ядро делится несколько раз с образованием 4—8 гаплоидных ядер, вокруг которых аккумулируется резервный материал (цитоплазма) и образуется общая стенка. Это и есть аск (греч. мешок), содержащий несколько аскоспор (рис. 7).

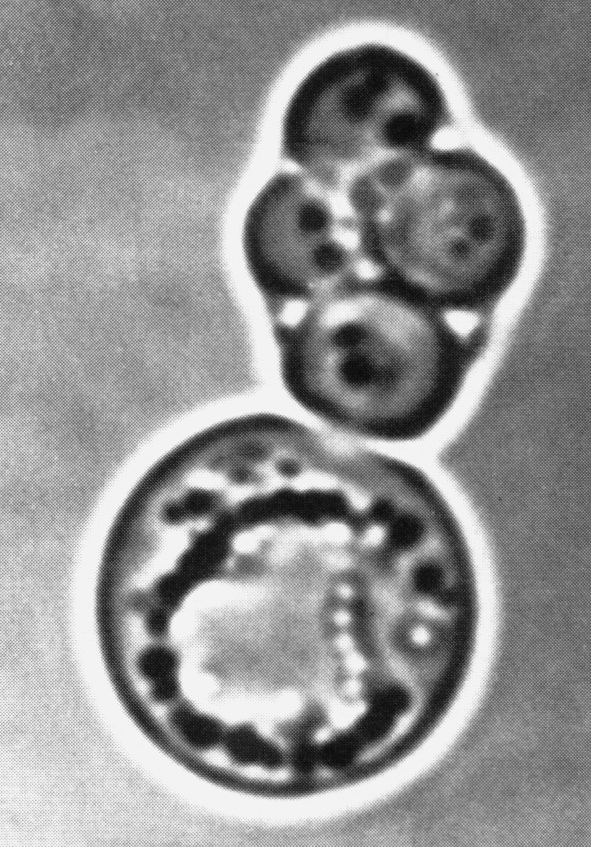
У плесневых аскомицетов аскоспоры образуются внутри специализированных гиф (асков), которые вырастают на основе зиготы, образующейся при слиянии ядер мужской и женской гамет (рис. 8, а). Из возбудителей микозов человека к аскомицетам относятся Piedra hortai (черная пьедра — разновидность трихоспории), Pseudallescheria boydii (мицетома), дерматофиты, Histoplasma capsulatum (гистоплазмоз) и Blastomyces dermatidis (бластомикоз).


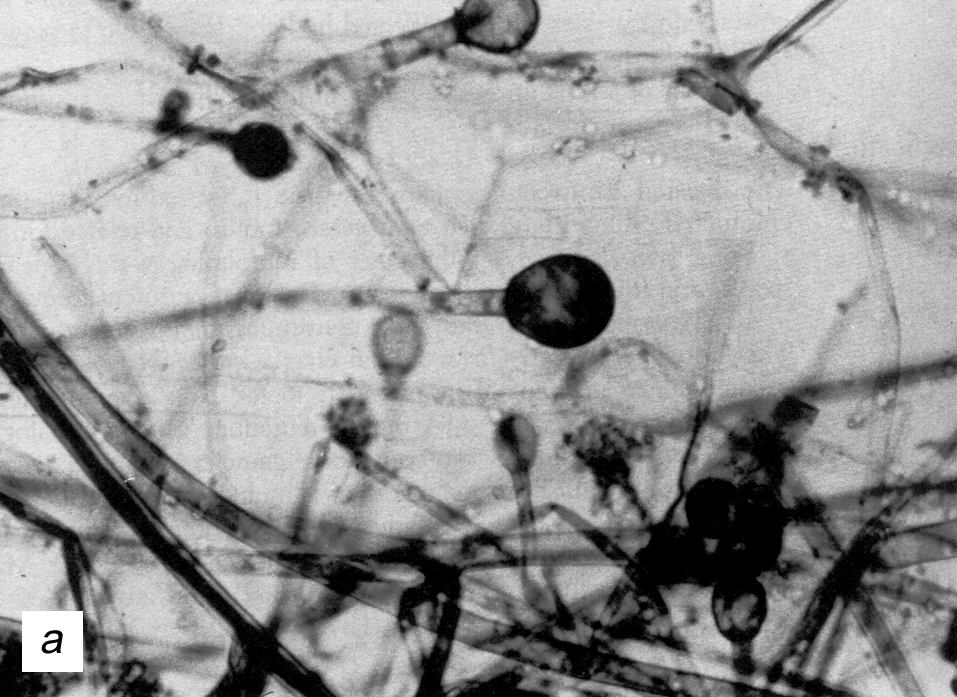
Рис. 8. Последовательные стадии образования половых спор у аскомицетов (а), базидиомицетов (б) и зигомицетов (в)
Базидиоспоры. Определяют принадлежность к базидиомицетам. Начальные события не отличаются от образования аскоспор. После слияния ядер зигота увеличивается в размерах, образуя булавовидную клетку — базидию (греч. basidion, уменьш. basis — основание). После мейоза ядро проходит через ряд митотических делений, давая несколько гаплоидных ядер. Дальнейший процесс протекает своеобразно. Споры образуются не внутри базидии, а на ее конце. Здесь возникают выросты (стеригмы), в которые переходят ядра. Вместе с цитоплазмой ядра отграничиваются перегородкой, образуя базидиоспоры, которые легко отрываются от основания — стеригм (рис. 8, б). К базидиомицетам относятся шляпочные грибы и единичные возбудители микозов (например, Cryptococcus neoformans).
Зигоспоры. Зигомицеты представлены в медицинской микологии несколькими видами родов Mucor и Rhizopus. Зигоспоры образуются путем слияния соседних гиф одного или разных мицелиев. Объединяющиеся гифы (они соответствуют мужской и женской гаметам) морфологически одинаковы, поэтому такой процесс спорообразования называют изогамией. Зигоспоры отличаются крупными размерами (30—50 мкм в диаметре) и их созревание примерно на неделю запаздывает от появления вегетативных спор (см. выше).
Морфологическая характеристика половых спор редко используется в качестве дифференциального признака в практических лабораториях, тем более что большинство возбудителей микозов до сих пор числятся среди дейтеромицетов.
Экология
Подавляющее большинство грибов являются сапрофитами. Это означает, что они живут за счет мертвого органического материала, т.е. органических остатков растений и животных. Грибы играют большую роль в экологическом балансе биосферы, разрушая сложные соединения (например, целлюлозу, кератин, пектин), которые плохо метаболизируются сообществом бактерий. Грибы годами сохраняют жизнеспособность при замораживании, а некоторые из них продолжают расти даже при минусовых температурах. Сапрофитные грибы лучше развиваются при комнатной температуре и заметно отстают в росте (или даже совсем не растут) при 37°С.
Но среди грибов есть и паразиты. В большинстве — это фитопатогенные виды, и лишь немногие агрессивны против человека и животных. Как возбудители биокоррозий грибы (особенно плесени) наносят большой экономический ущерб, поражая объекты сельского хозяйства и промышленности. Впрочем, это с лихвой компенсируется полезной деятельностью грибов и их широким использованием в биотехнологии, в том числе для получения ценных лекарственных препаратов (антибиотики, стероидные гормоны и пр.).
Большинство возбудителей микозов относится к термотолерантным сапрофитам, для которых заражение человека и животных — случайный, необязательный эпизод в их жизненных циклах. Как правило, инфекция не получает распространения среди людей (животных), так как в таких случаях возбудитель попадает в ситуацию, которую принято называть экологическим тупиком.
Из единичных представителей фунгальной флоры человека наиболее известны дрожжеподобные грибы рода Candida, в основном C. albicans. Они вегетируют в кишечнике, ротовой полости, влагалище и на коже здоровых людей, создавая прецедент для оппортунистических инфекций. Нередко с кожи здоровых людей выделяется Pityrosporum/Malassezia furfur. Это липофильные дрожжи, которые растут за счет липидов, секретируемых сальными железами кожи. По неизвестным причинам они могут усиленно размножаться в роговом слое эпидермиса, вызывая разноцветный/отрубевидный лишай или себорейный дерматит.
Допускается бессимптомное носительство на коже одного из возбудителей трихоспории (дрожжи Trichosporon beiglii — белая пьедра), некоторых дерматофитов (например, возбудителя эпидермофитии стоп — Trichophyton mentagrophytes var. interdigitale) и Pneumocystis carinii (возбудитель оппортунистической инфекции у больных СПИДом).
Позиция P. сarinii среди грибов поддерживается главным образом на основании гомологии рибосомальных РНК; во всем остальном они больше похожи на простейшие, в частности, не культивируются in vitrо и не чувствительны к антифунгальным препаратам. Если убиквитарность пневмоцистной инфекции не вызывает сомнений (большинство детей уже имеют антитела к P. carinii), то облигатное носительство ее признается не всеми. Вполне вероятно, что пневмоцистная инфекция в норме кратковременна, и подкрепление иммунитета обеспечивается периодическим реинфицированием из внешней среды. Источники инфекции гипотетичны.
Не все грибы одинаково распространены в природе. Кроме убиквитарных видов есть грибы, которые встречаются в определенных, чаще тропических регионах Земного шара. Это характерно и для ряда патогенных грибов, что определяет эндемичность вызываемых ими заболеваний (южно-американский бластомикоз, северо-американский бластомикоз, мадуромикоз, пенициллез, вызываемый P. maneffrei и др.). По неизвестным причинам патогенность грибов может не соответствовать их глобальному распространению. Так, например, обстоит дело с гистоплазмозом: он встречается преимущественно в некоторых регионах США (долины рек Миссисипи и Огайо), в Центральной и Южной Америке, хотя возбудитель (Histoplasma capsulatum) убиквитарен.
Наиболее сложна эпидемиология дерматофитозов — фунгальных (грибковых) поражений кожи и ее придатков, вызываемых дерматофитами, к которым относятся более 40 видов родов Microsporum, Trichophyton и Epidermophyton. Эколого-патогенетической основой является способность дерматофитов утилизировать кератин — базисный белок рогового слоя эпидермиса (кератиноцитов), ногтей, волос, копыт и рогов. Дерматофиты имеют массу потенциальных экологических ниш, так как живой и мертвый кератин широко распространены в природе. Действительно, часть из них имеет почвенный резервуар, но большинство ведут паразитический образ жизни и, по-видимому, лишены сапрофитической фазы. Многие дерматофитозы проявляют отчетливую зависимость от пола и возраста, некоторые имеют ограниченное географическое распространение (это прежде всего относится к микозам Южной Америки, Африки, Азии). Известны антропофильные дерматофиты, которые строго адаптированы к человеку и не инфицируют животных (например, Microsporum audounii — микроспория, Trichophyton rubrum — трихофития, Trichophyton schoenleinii — парша). Другие в качестве естественных хозяев используют животных (например, кошек и собак) и от них могут передаваться человеку. Такие грибы называются зоофильными (например, Microsporum canis). Наконец, заражение некоторыми (геофильными) дерматофитами происходит из почвы (например, Microsporum gypseum).
Микозы
Патология
Инвазия грибов в ткани сопровождается воспалительной реакцией, в которой доминируют гранулематозный (макрофагзависимый) или пиогенный (нейтрофилзависимый) компоненты. Первый обычно захватывает лидерство, а стабилизация пиогенности часто отражает присоединение бактериальной инфекции (например, при дерматофитозах). Возбудители микозов не продуцируют собственных токсинов, и все токсические эффекты опосредованы через медиаторы воспалительных реакций. В патогенезе могут быть задействованы иммунологические (Т-зависимые) механизмы. Усиливая гранулематозное воспаление, они переводят его в ранг иммунопатологических реакций, которые содействуют повреждению тканей. В целом микозы имеют тенденцию к хроническому течению, хотя нередко и начинаются остро.
В классификации микозов ориентируются на два опорных признака — локализацию процесса и агрессивность возбудителя против нормального и ослабленного хозяина.
Поверхностные микозы. Развитие грибов происходит на поверхности эпителиальных тканей (эпидермис, волосы, ногти) без заметной пенетрации в живую ткань. Иммунный ответ и воспалительная реакции отсутствуют. Экологически такие микозы сопоставимы с эпифитным и эпилитным (от греч. lithos — камень) ростом, т.е. с поверхностной колонизацией растений, древесины, камней, эпидермиса, которая происходит практически без использования поддерживающего субстрата (колонизация эпидермиса, по-видимому, сопровождается утилизацией продуктов, экскретируемых кожей). Примерами являются разноцветный (отрубевидный) лишай (возбудитель — Pityrosporum/Malassezia furfur, биовар orbiculare) и себорейный дерматит (P. furfur, биовар ovale) (рис. 9).


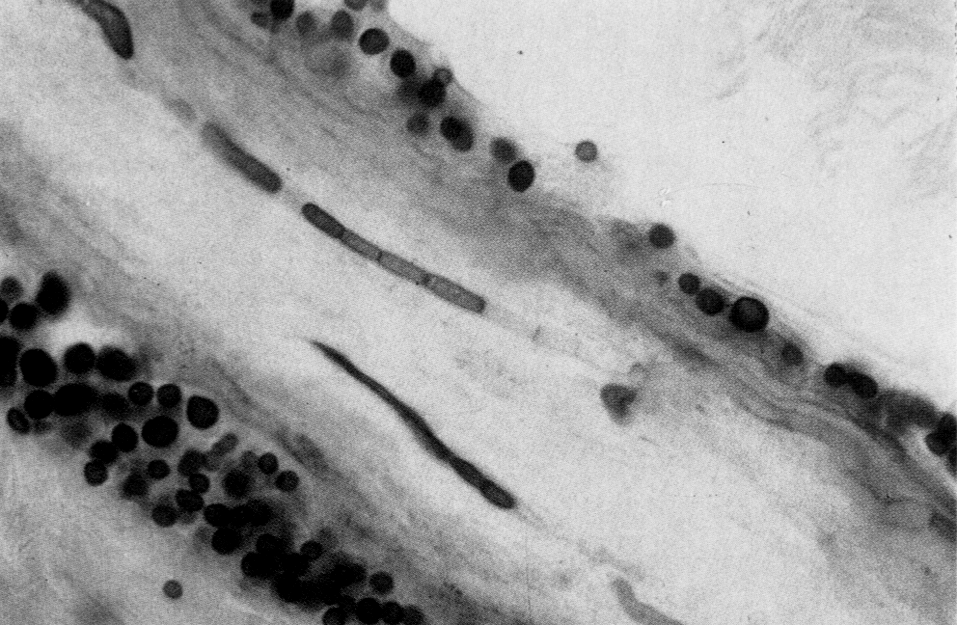

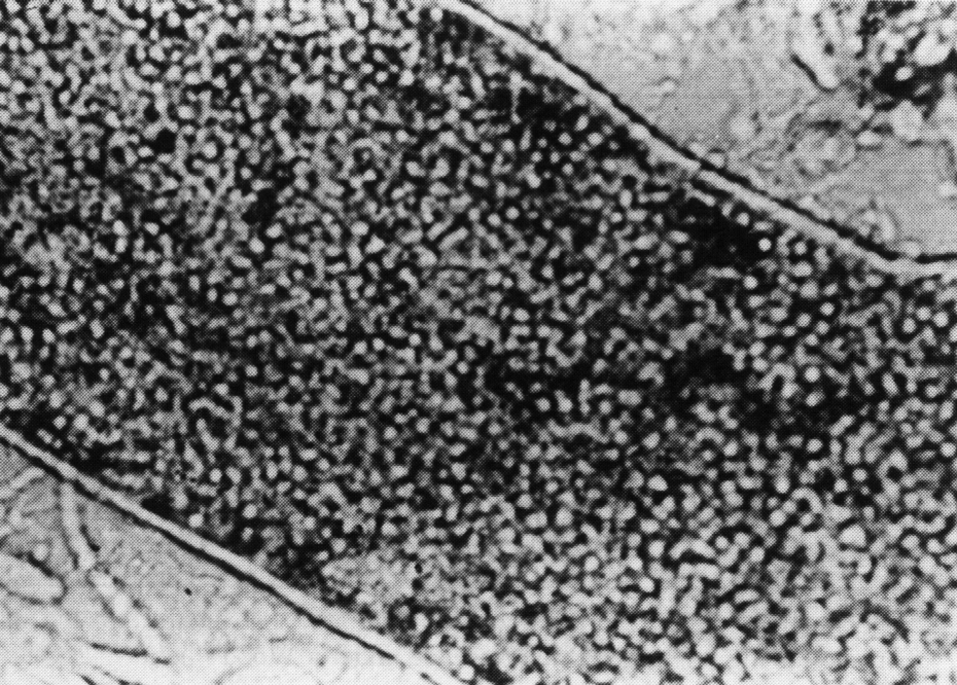
Кожные микозы. В большинстве случаев причиной являются дерматофиты — представители родов Microsporum, Trichophyton и Epidermophyton. Вызываемые ими микозы называются дерматофитозами. Инфекция ограничена роговым слоем эпидермиса (stratum corneum), поражением ногтей и волос (рис. 10, 11), но основная симптоматика обусловлена воспалительной реакцией в эпидермисе, дерме и волосяных фолликулах.
Раздражающее действие фунгальных метаболитов запускает острое воспаление, а проникновение антигенов в эпидермис возбуждает иммунный ответ с развитием гиперчувствительности замедленного типа. Это усугубляет патологический процесс, переводя его на рельсы иммунного воспаления. Интенсивность воспалительной реакции коррелирует с уровнем гиперчувствительности кожи к экстрактам из дерматофитов.
Поражения кожи и ногтей (онихомикоз) могут вызывать Candida и (крайне редко) недерматофитные плесени (различные виды Aspergillus, Penicillium, Alternaria, Fusarium и др.). Это всегда вторичные (оппортунистические) инфекции, сопряженные с ослаблением резистентности организма. Впрочем, и дерматофитии нередко наслаиваются на преморбидный фон. Например, частота микозов стоп с поражением ногтей почти в 3 раза выше у больных сахарным диабетом, чем в общей популяции (ростстимулирующий эффект глюкозы!?).
Подкожные микозы. Возникают при травматическом заносе спор грибов-сапрофитов в подкожную ткань (например, с занозами). Зарождается хроническая воспалительная реакция (гранулема, иногда с пиогенным компонентом), которая распространяется на кожу (дерму и эпидермис), но обычно не выходит за пределы первичной очаговости. Большинство подкожных микозов встречается в странах с тропическим климатом. Повсеместное распространение имеет лишь споротрихоз (Sporothrix schenckii).
Системные (глубокие) микозы. Сюда относятся самые тяжелые, часто фатальные формы микозов. Их принято делить на первичные и оппортунистические (вторичные). Возбудители первичных микозов поражают здоровых людей, грибы-оппортунисты отличаются условной патогенностью и опасны только на фоне ослабления иммунитета.
При первичных микозах входными воротами для спор служат легкие, откуда болезнь распространяется на другие участки тела. В очагах колонизации развиваются туберкулоидные гранулемы, иногда с нагноением. К наиболее известным микозам этой группы относятся гистоплазмоз (Histoplasma capsulatum), бластомикоз (Blastomyces dermatidis), кокцидиомикоз (Coccidioides immitis) и паракокцидиомикоз (Paracoccidioides brasiliens). Они имеют ограниченное географическое распространение (исключение с некоторыми оговорками составляет гистоплазмоз), их возбудители являются сапрофитами и характеризуются температурозависимым диморфизмом. Убиквитарность спор в эндемичных зонах делает заражение вполне обычным явлением. Об этом судят по иммуноэпидемиологическим данным (наличие антител и/или положительных кожных тестов с антигенами грибов), рентгенологическим и аутопсийным находкам (следы реакций, закончившихся излечением). Инфицирование, как правило, завершается субклиническим конфликтом или малым, самокупирующимся заболеванием. Лишь изредка заражение переходит в тяжелый, часто фатальный процесс, свидетельствуя о том, что для проявления вирулентности необходима солидная инфицирующая дозировка и, возможно, генетическая предрасположенность хозяина.
Что касается возбудителей оппортунистических микозов, то они бессильны против нормальной физиологии, и лишь гипорезистентность предрасполагает к реализации их болезнетворности. Группы риска составляют больные диабетом, ожогами, травмами, злокачественными опухолями, с пересадками органов (они получают интенсивную цитостатическую и иммуносупрессивную терапию), недоношенные новорожденные, лица, нуждающиеся в парентеральном питании или длительной внутрисосудистой катетеризации, наркоманы (нестерильные инъекции и иммунодепрессивное действие опиатов).
Предрасполагающим фактором является злоупотребление антибиотиками, особенно широкого спектра действия. Угнетая нормальную микрофлору, они ослабляют один из важных механизмов колонизационной резистентности слизистых оболочек, склоняя к бесконтрольному размножению таких грибов, как Candida, аспергиллы и др. Парадокс, но именно блестящие достижения медицины спровоцировали рост и опасные последствия фунгальных инфекций. Серьезной поддержкой для грибов-оппортунистов стало появление в начале 1980-х гг. СПИДа — ретровирусного синдрома приобретенного иммунодефицита.
Большинство возбудителей оппортунистических микозов относятся к сапрофитам, которые инфицируют человека через дыхательные пути или (редко) пищеварительный тракт. К наиболее известным заболеваниям этой группы относятся криптококкоз (возбудитель — Cryptococcus neoformans), аспергиллез (обычно A. fumigatus), зигомикоз (некоторые виды Rhizopus, Mucor и др.) и (очень редко) пенициллез (некоторые виды Penicillium). Впрочем, спектр такого рода микозов постоянно расширяется, прежде всего в связи с распространением СПИДа. Некоторые из них географически ограничены, встречаясь главным образом в странах тропического региона (например, пенициллез, вызываемый Penicillium marneffei — диморфной плесенью, распространенной в Юго-Восточной Азии).
Однако в количественном отношении все перечисленные заболевания, безусловно, уступают кандидозу — едва ли не единственному оппортунистическому микозу эндогенной природы. Его вызывают более десятка видов дрожжеподобных грибов рода Candida, но абсолютным лидером остается C. albicans (за ним следуют C. tropicalis, C. parapsilosis, C. krusei, C. lusitaniae и C. (Torulopsis) glabrata). Кандидоз — самый распространенный из системных микозов человека, так как носительство C. albicans носит практически поголовный характер и любое серьезное нарушение местного или общего иммунитета может спровоцировать их активацию. Возможно и экзогенное инфицирование, особенно в госпитальных условиях, что более типично для минорных (менее распространенных) видов кандид.
Кандидоз проявляется необычайно широким спектром клинических патологий — от ограниченных поражений кожи и слизистых оболочек (молочница, вульвовагинит) до системного поражения мукозального тракта и гематогенных метастазов во внутренние органы (пневмония, менингит, эндокардит, остеомиелит, артрит и пр.), от поражения ослабленных новорожденных до тяжелых заболеваний иммунокомпрометированных взрослых людей. Кстати, концепция о «скомпрометированном больном» была сформулирована французскими педиатрами Труссо и Парро в 1868—1869 гг. в связи с наблюдениями о диссеминированном кандидозе новорожденных с атрепсией.
Патогенез
Наиболее общим признаком фунгальных видов, потенциально опасных для человека, является термотолерантность, т.е. способность активно размножаться при температуре тела хозяина. Это исключает опасность огромного количества грибов-сапрофитов, которые не способны преодолеть этот рубеж видового иммунитета. Например, из всех видов аспергилл лучше всего приспособлен A. fumigatus, который хорошо растет в диапазоне 20—50°С. К базисным факторам вирулентности относится и температурозависимый диморфизм. Он обеспечивает трансформацию грибов в инвазивные формы, отражая высокую адаптивную изменчивость, основанную на механизме фенотипических переключений.
Возбудители микозов не продуцируют специфических токсинов, и патогенез фунгальных инфекций сводится к воспалительным процессам, возбуждаемым микробной инвазией. Для возбудителей системных микозов характерна инвазия в стенки сосудов с развитием тромбоза, ишемии и тканевого некроза, открывающего путь инфекции в глубжележащие ткани. Ослабление эффекторного потенциала иммунитета (нарушение Т-зависимой активации макрофагов, нейтропения) ведет к опережающему размножению грибов, вынуждая к количественному усилению воспалительных реакций за счет малоэффективной мобилизации эффекторов воспаления.
Но грибы пользуются не только пассивными дефектами хозяина. Некоторые из них активно блокируют реакции иммунитета, подавляя функции Т-лимфоцитов, макрофагов, нейтрофилов, комплемента (глиотоксин A. fumigatus, маннан C. albicans и др.). Классическим примером антифагоцитарной устойчивости являются инкапсулированные криптококки, но и другие грибы создают немало проблем для фагоцитов. Например, гифы, образующиеся после прорастания спор, слишком велики для поглощения и могут быть убиты только извне — биооксидантами, секретируемыми нейтрофилами и макрофагами. Некоторые грибы (например, C. albicans, A. fumigatus) сорбируют фибриноген и другие белки, защищая себя от фагоцитоза псевдокапсулой.
Грибы располагают широким арсеналом адгезивных факторов, которые обеспечивают закрепление на различных тканевых и искусственных субстратах. Рост грибов, следующий за адгезией, сопровождается продукцией гидролитических ферментов (протеазы, фосфолипазы), которые могут повреждать тканевые барьеры, содействуя пенетрации. Их концентрация выше на концах растущих гиф, что объясняет усиление инвазивности при прорастании спор, например при образовании ростковых трубок у C. albicans (рис. 12).

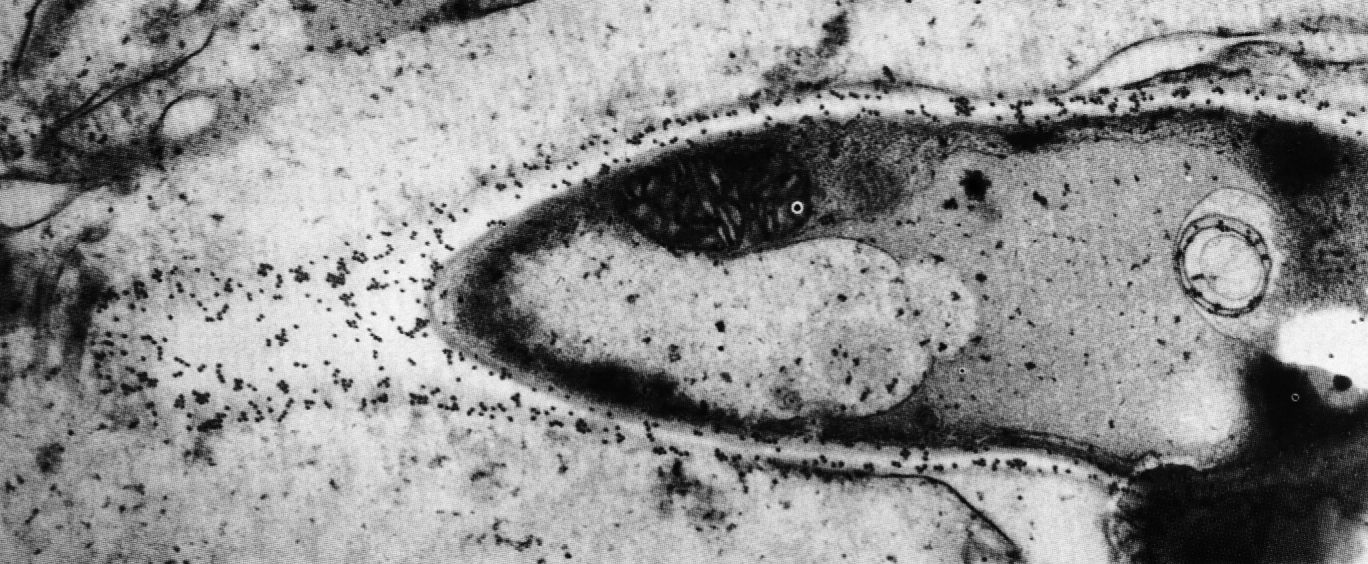
Имеет значение и такой простой фактор, как размер спор (конидий). У всех возбудителей системных микозов он не превышает 5 мкм, обеспечивая проникновение в альвеолы при ингаляционном заражении.
Иммунитет
Фунгальные инфекции (за исключением поверхностных, неинвазивных микозов) сопровождаются широким спектром иммунных реакций, которые обеспечивают подключение различных эффекторов иммунитета. Главную роль играют два механизма:
- Т-зависимые реакции макрофагов, действующие в рамках гранулематозного воспаления;
- нейтрофилы, поддерживаемые антителами и комплементом.
О значении эффекторов Т-клеточного иммунитета свидетельствуют патогистологическая картина очагов поражения, развитие гиперчувствительности замедленного типа к фунгальным антигенам и гипорезистентность к возбудителям оппортунистических микозов у лиц с ослаблением Т-клеточного звена иммунитета. Внутривенное введение дериватов клеточных стенок грибов (например, зимозана) вызывает развитие множественных гранулем в органах, богатых ретикулярной тканью. Их эволюция, поддержанная сенсибилизированными CD4 Т-лимфоцитами и исходно нацеленная на уничтожение возбудителя, иногда выходит за рамки компенсаторных реакций, превращаясь в инструмент патогенеза. Парадоксально, но этому способствует не только гиперреактивность, но и ослабление СD4 Т-лимфоцитов. В этом случае макрофаги лишаются активирующих сигналов Т(Тh1)-цитокинов, что резко снижает коэффициент полезного действия гранулем, провоцируя количественное усиление процесса без должной эффекторной отдачи.
Пиогенный компонент (не связанный с вторичной бактериальной инфекцией), который нередко присоединяется к гранулематозным реакциям, отражает подключение нейтрофилзависимого механизма защиты, усиливаемого антителами и комплементом. Ослабление любого из трех элементов (фагоциты—антитела—комплемент) снижает коэфициент полезного действия реакции, предрасполагая к инвазивным осложнениям. Не случайно больные с нейтропенией составляют группу риска по развитию генерализованных форм оппортунистических микозов. Этим, в частности, можно объяснить редкие случаи системного кандидоза, аспергиллеза и зигомикоза у больных СПИДом, когда фагоцитарный механизм антиинвазивной защиты страдает не столь заметно, как CD4 Т-клеточный иммунитет. Напротив, кандидоз слизистых оболочек принадлежит к категории СПИД-индикаторных инфекций.
Следует учитывать и степень колонизационной резистентности инфицируемых тканей. С этим, например, могут быть связаны физиологические (возрастные и половые) особенности чувствительности к дерматофитам и другим патогенным грибам. Предрасположенность к кожно-слизистому кандидозу у больных тяжелым диабетом может быть следствием гипергликемии, содействующей размножению кандид. Активация C. albicans при антибиотикозависимых дисбактериозах объясняется снижением конкурентной защиты слизистых оболочек, обусловленной бактериальной нормофлорой. То же самое замечено и для других оппортунистических микозов, например аспергиллеза. Не исключено, что некоторые ятрогенные воздействия (цитостатики, иммунодепрессанты, стероиды) негативно влияют не только на систему иммунитета, но и на колонизационную резистентность эпителиоцитов, содействуя тем самым микробной инвазии. Изучение этого вопроса может внести коррективы в концепцию о компрометированном хозяине, в частности в представления о патогенезе фунгальных инфекций.
Антифунгальные препараты
Наиболее очевидными мишенями для избирательного воздействия на грибы являются плазматическая мембрана и клеточная стенка. В первом случае речь идет об эргостероле и его синтезе (присутствие эргостерола наиболее радикально отличает грибы от высших эукариот), во втором — о выключении синтеза полисахаридных структур, уникальных для клеточной стенки грибов. Известны и другие молекулярные мишени, которые уже использовались как объекты антифунгальной терапии (гризефульвин — ингибитор микротубулярного аппарата фунгальных клеток, флюцитозин — ингибитор синтеза ДНК/РНК) или рассматриваются среди ее потенциальных кандидатов. К ним относятся ингибиторы фактора элонгации-2, топоизомеразы и ряда других ферментов, в том числе специфичных для некоторых грибов (например, фенилоксидазы и ферментов синтеза капсулы Cryptococcus neoformans — возбудителя криптококкоза).
Ингибиторы плазматической мембраны представлены полиенами, производными азолов, аллиламинами и морфолинами. Полиены относятся к макролидным антибиотикам с ненасыщенными диеновыми связями (метаболиты нескольких видов стрептомицетов). Они открыты в начале 1950-х гг. и были первыми антифунгальными препаратами направленного действия. Избирательность эффекта полиенов основана на преимущественном связывании с эргостеролом (слабее — с другими стеролами), что ведет к осмотической нестабильности и нарушению целостности мембраны. Полиены характеризуются одинотипным и широким спектром антифунгальной активности, но токсичны для хозяина. Единственным препаратом, рекомендованным для системного (внутривенного) применения, является амфотерицин В (продукт Streptomyces nodosus). Из остальных полиенов значение сохраняет лишь нистатин, который (не очень убедительно) активен при кандидозе слизистых оболочек.
Новую эру в антифунгальной терапии открыли производные азола (конец 1960-х гг.). Их эффект объясняется блокадой синтеза эргостерола (на уровне цитохром Р-450-зависимой ланостерол-деметилазы). Действие азолов проявляется через несколько генераций грибов, а потому заметно отстает от полиенов. Однако азолы отличаются более высокой избирательностью и, вследствие этого, меньшей токсичностью. На смену имидазолам (миконазол, кетокиназол, клотримазол, эконазол) пришло поколение триазолов (флуконазол, итраконазол, тетраконазол и др.) с улучшенной фармакокинетикой и переносимостью. Они различаются по спектру антифунгальной активности и фармакологическим параметрам, что позволяет варьировать их применение в зависимости от возбудителя и клинической формы микозов. Например, флуконазол высокоактивен против дрожжеподобных грибов, но в 10—100 раз слабее итраконазола при большинстве других микозов. В сочетании с хорошей проницаемостью в центральную нервную систему это определяет его преимущество при лечении кандидозного и криптококкозного менингитов. Получено много новых триазолов, обладающих рядом достоинств по сравнению с уже известными препаратами. Дело не только в фармакодинамике, но и в воздействии на резистентные штаммы, эволюционирующие под влиянием химиотерапии.
Производные азола не решили всех проблем антифунгальной терапии. Например, зигомицеты не чувствительны к азольным препаратам, хотя содержат в своих мембранах эргостерол. Главным средством в этом случае (как и при лечении ряда других микозов) остается амфотерицин В.
Подобно азольным производным, дериваты морфолина (аморолфин) и аллиламины (нафтифин, тербинафин) подавляют синтез эргостерола, но действуют на других уровнях эргостерольного каскада. Их применение ограничено местным (аморолфин, нафтифин) и пероральным (тербинафин) лечением дерматофитозов, чему способствуют высокая липофильность (особенно аллиламинов) и аккумуляция в ногтевом ложе (важно для лечения онихомикозов). Аморолфин с успехом применялся при вагинальном кандидозе.
Внедрение ингибиторов синтеза клеточной стенки только начинается, но видится как обнадеживающая перспектива. Среди них природные и полусинтетические липопептиды и липосахариды, нацеленные на подавление синтеза глюканов (эхинокандины, пневмокандины, папулокандины, малюнокандины), хитина (никкомицины) и других компонентов клеточной стенки грибов (прадимицин).
Резистентные штаммы появились вскоре после внедрения полиенов, но реальные опасения возникли в связи с широким использованием азольных производных. Кроме отбора резистентных клонов, возникающих в результате случайных мутаций, этому способствует фенотипическая вариабельность (и соответственно экологическая пластичность) грибов, позволяющая выживать в неблагоприятных условиях. Лучше всего это изучено на модели так называемого феномена переключения у C. albicans. Он обусловлен специальным механизмом перестройки генетического аппарата, который обеспечивает мозаичность фенотипических вариантов. Это содействует отбору клеток, наиболее приспособленных к конкретной экологии, в том числе резистентных к антифунгальным агентам. Стратегия адаптивной резистентности, основанная на селекции и эволюции случайных мутантов, находит здесь мощную тактическую поддержку, базирующуюся на дифференцированной экспрессии преформированных генов.
Механизмы устойчивости к азолам определяются снижением аффинности фермента-мишени (ланостерол-деметилазы) к азольным производным, утратой активности фермента-мишени, гиперпродукцией ланостерол-деметилазы, снижением проницаемости клеток для антифунгальных агентов, заменой эргостерола на другие стеролы (этот механизм действует и против полиенов) и ускоренным выведением агентов из клеток. Обнадеживает то, что резистентность даже против препаратов одного класса не абсолютна. Например, изоляты кандид, устойчивые к флуконазолу, могут сохранять чувствительность к другим производным азола.
Диагностика
Для выявления фунгальных инфекций большое значение имеет обычная микроскопия исследуемого материала. Фиксированные препараты можно окрасить практически любыми методами, но часто исследуют нативные (влажные) препараты. Это особенно важно при плесневых микозах, когда необходимо получить верное представление о морфологии мицелия — опорного признака видовой дифференциации плесневых грибов. Анализируемый материал подвергают обработке крепкой щелочью; растворяя кератин и другие тканевые компоненты, он не влияет на структурные элементы грибов. В диагностике глубоких микозов важную роль играет патогистологическая характеристика пораженных тканей.
Для уточнения прибегают к культуральному анализу. Инкубацию проводят при 37 и 24оС (комнатной температуре), что позволяет выявить диморфизм и ускорить рост сапрофитных грибов, которые медленно размножаются при температуре тела. В целом культивирование не представляет трудностей. Исключением служат первично-патогенные возбудители системных микозов, которые, хотя и не требуют особых условий для культивирования, но растут очень медленно (несколько недель). Грибы любят высокое содержание углеводов и подкисленную среду, задерживающую размножение бактерий. Это учитывают при изготовлении питательных сред. Кроме того, в них часто добавляют антибиотики, которые, подавляя рост бактерий, не влияют на развитие грибов. Для выделения медленно растущих грибов в среды вносят циклогексимид (ингибитор белкового синтеза). Он действует на быстро делящиеся клетки, но малотоксичен в отношении медленно растущих грибов. Культуры идентифицируют по типичным признакам. При изучении плесневых грибов опираются на морфологию мицелиальных структур (прежде всего конидиогенные гифы и конидии), анализ дрожжеподобных грибов (кандид) дополняют определением ферментативной активности.
Образование антител при системных микозах служит основанием для серологической диагностики гистоплазмоза, бластомикоза и пр. Наиболее достоверным является нарастание титров антител (сероконверсия) в ходе заболевания, так как небольшое количество антител может быть результатом прежних субклинических контактов или здорового носительства (Candida). Предложены препараты для кожно-аллергических тестов, но их надежность невелика.
Новые подходы к лабораторной диагностике микозов основаны на выявлении в сыворотке и других субстратах фунгальных антигенов (например, маннана при кандидозе и галактоманнана при аспергиллезе), специфических метаболитов (d-маннитол, щавелевая кислота при аспергиллезе) и генетических маркеров (полимеразная цепная реакция).
Микоаллергозы
Cпоры грибов составляют важнейший элемент так называемых аэроаллергенов — иммунологически активных продуктов, которые поступают в атмосферу из биологических или промышленно-бытовых источников и, проникая через респираторный тракт, вызывают сенсибилизацию организма. Особенно характерны заболевания респираторного тракта (бронхиальная астма, альвеолиты, риниты), которые формируются на основе IgE-зависимых реакций и иммунокомплексной патологии.
В количественном выражении споры грибов значительно превосходят содержание в воздухе таких классических аллергенов, как зерна пыльцы растений. К примеру, соотношение между спорами Cladosporium и гранулами пыльцы составляет 1000:1. Важным источником фунгальных спор служит домашняя пыль: в 1 г различных образцов пыли содержится от 22 тыс. до 11250 тыс. жизнеспособных спор плесневых грибов.
К наиболее важным источникам аллергенов относится так называемая большая четверка — различные виды Aspergillus, Penicillium, Alternaria, Cladosporium. Кроме них имеют значение и другие грибы: Mucor, Rhizopus, Fusarium, Epicoccum и пр.
Клинико-эпидемиологической особенностью фунгальной аллергии, которая отличает ее от пыльцевой сенсибилизации, является отсутствие выраженной сезонности, связанной с периодами цветения растений. Споры грибов распространены постоянно и повсеместно, они присутствуют не только в наружной атмосфере, но насыщают жилые и служебные помещения, поддерживая аллергизацию и провоцируя обострения. Замечено, что в жилищах больных бронхиальной астмой фунгальных спор может быть в несколько раз больше, чем в других помещениях. Накопление спор зависит от температуры, влажности, освещенности, степени загрязнения, воздушных потоков. Этим объясняются значительные особенности содержания спор в пыли различных бытовых и производственных помещений. Например, на текстильных предпиятиях, производящих хлопковые и шерстяные ткани, преобладают споры Aspergillus, Penicillium, Fusarium. На кожевенных производствах доминируют Mucor, Rhizopus, Nigrospora. На мукомольнях к ним добавляются Alternaria и др. Вспышки аллергических заболеваний отмечались на предприятиях, занятых фунгальной биотехнологией (производство антибиотиков, кормовых дрожжей и др.). Это определяет конкретные задачи по профилактике фунгальной сенсибилизации и аллергических заболеваний.
Ряд грибов, выступающих в роли аллергенов человека, патогенны для растений, вызывая у них заболевания в виде гнилей и плесеней (cм. таблицу).
Фитопатогенность некоторых грибов, способных аллергизировать человека
| Грибы | Растение | Заболевания |
| Альтернария (A. brassicae и др.) | Картофель | Бурая пятнистость листьев |
| Морковь | Черная гниль | |
| Капуста | Черная пятнистость листьев | |
| Табак | Черная и серая гниль рассады | |
| Кладоспориум (C. fulvum) | Помидоры, персики | Бурая пятнистость листьев |
| Гельминтоспориум | Ячмень, пшеница |
Полосатая, сетчатая, бурая пятнистость |
| Ионилия | Хлопчатник | Шоколадная гниль |
| Вишня, слива, груша | Серая плодовая гниль | |
| Ризопус (R. nigricans) | Ягоды | Черная плесень |
| Хлопчатник | Черная гниль | |
| Мукор (M. ingricus) | Яблоки, слива | Гниль |
| Аспергиллус (A. niger) | Винная ягода, финики | Ржавчина, черная плесень |
| Пенициллиум (P. crustaceum и др.) |
Хлопчатник | Черная гниль |
| Цитрусовые | Голубая или зеленая плесень | |
| Фузариум (F. solani и др.) | Помидоры | Белая гниль |
| Картофель | Водянистая гниль | |
| Злаки (овес, пшеница, рожь) | «Выпаривание» злаков (снежная плесень) |
Микотоксикозы
Микотоксикозами называются отравления микотоксинами — ядовитыми метаболитами плесневых грибов. С этим чаще сталкиваются ветеринарные врачи, но и для человека микотоксикозы представляют реальную опасность. Началом изучения токсинообразующих грибов можно считать середину XIX в., когда была установлена принадлежность к грибам рожков спорыньи — Claviceps purpurea. Они поражают рожь и некоторые другие виды зерновых культур, вызывая у человека заболевание, известное как эрготизм (от старофранцузского argot — петушиная спора, по виду рожков спорыньи на пораженных колосьях). В 1941—1945 гг. была доказана фунгальная природа алиментарной токсической алейкии человека (Fusarium sporotrichiella).
Статус самостоятельной науки микотоксинология получила после открытия афлатоксинов (1960 г.). Это соединения типа фурокумаринов, которые продуцируются широко распространенными плесневыми грибами рода Aspergillus. К счастью, такой способностью обладают лишь немногие штаммы двух видов аспергилл: Aspergillus flavus (отсюда название — афлатоксины) и Aspergillus parasiticus. Известны и другие микотоксины: охратоксины (Aspergillus ochraceus), метаболиты родов Penicillium, Alternaria и пр. Всего известно более 200 видов плесеней, продуцирующих примерно сотню метаболитов, токсичных для разных биологических систем. Например, к афлатоксинам чувствительны большинство млекопитающих, различные виды рыб, насекомых, растений и микроорганизмов. Этому не стоит удивляться. Антибиотики, продуцируемые грибами (пенициллин, цефалоспорин и др.), которые широко используются в лечении инфекционных заболеваний, по сути те же микотоксины, но избирательно действующие на бактерии.
Все микотоксины — вещества небелковой природы, с низкой молекулярной массой (менее 1000 Да), термостабильны (выдерживают длительное кипячение), лишены антигенных свойств. Это вторичные метаболиты, образующиеся в цепи последовательных ферментативных реакций из относительно небольшого числа промежуточных продуктов основного метаболизма. Микотоксины принадлежат к наиболее сильнодействующим клеточным ядам. Особенно активен афлатоксин В1 — самый сильный (для человека и животных) из биологических ядов. Он вызывает некротические изменения во многих внутренних органах, прежде всего в печени. Некоторые микотоксины обладают мутагенным и канцерогенным действиями. К примеру, в опытах на животных канцерогенность афлатоксина В1 в 1000 раз выше, чем эффект самых сильных химических канцерогенов.
Микотоксикозы обычно возникают после употребления продуктов, пораженных грибами. Например, эрготизмом и алиментарно-токсической лейкоплакией можно заболеть после употребления пищи, приготовленной из пораженных злаковых растений (хлеб, пшенная каша и пр.). Наиболее опасно перезимовавшее зерно, впитавшее ядовитые метаболиты паразитирующих на нем грибов. Известны примеры неалиментарного отравления микотоксинами — через кожные покровы и слизистые оболочки дыхательных путей. В таких случаях в анамнезе имеется, как правило, контакт с сырьем, обсемененным спорами грибов (хлопок, корма для скота, сырьевая пыль и пр.). Это казуистика, но она расширяет представления об эколого-патогенетических ресурсах грибов, которые раскрываются в нестандартных ситуациях.
* * *
Медицинская микология долго оставалась на положении Золушки. Практический интерес к фунгальным инфекциям был ограничен главным образом сферой дерматологии. Глубокие микозы не вызывали большого интереса, так как наблюдались редко и часто были географически ограничены. Сегодня такой стереотип уже не устраивает. Принципиальные изменения, произошедшие в медицине за последние десятилетия, серьезно повлияли на этиологическую структуру инфекционной патологии, сместив акцент в пользу заболеваний, вызываемых условно-патогенными микроорганизмами. В этой парадоксальной, но вполне объяснимой ситуации микозы стали заметным явлением. Некоторые из них уже заняли прочное место в госпитальной практике, войдя в число угрожающих оппортунистических инфекций. Медицинская микология переживает второе рождение, эволюционируя на основе современных достижений молекулярной биологии, биохимии и иммунологии. Подчиняясь традиционной (в целом оправданной) доминанте бактериологического и вирусологического мышления, не следует забывать еще об одном лике микробиологии — микологии. Иначе Золушка может обернуться злой, неуправляемой принцессой.
Послесловие
Мысль о написании этой книги возникла много лет назад как реакция на курьезные ситуации, с которыми автор столкнулся на студенческих экзаменах по микробиологии. Ограничусь одним примером. Говоря о сальмонеллах, мой юный коллега справедливо отметил их подвижность и интригующе добавил, что они располагают двадцатью четырьмя жгутиками. Удивившись такой точности, я спросил, может ли быть у сальмонелл двадцать три жгутика? «Может, но редко!». Псевдодеталей подобного рода я слышал немало. Они либо неверны, либо не требуются врачу для общей микробиологической культуры и понимания механизмов микробных болезней. Познакомившись с учебниками, я убедился, что они тоже грешат сходными «перлами» и, втягивая читателя в ненужные дебри, отвлекают от главного. Официальная критика, ограничиваясь замечаниями тактического плана, не касается стратегии изложения материала. Между тем, известны старые (но не устаревшие!) издания, авторы которых следовали принципу рационального отбора фактов для базисного курса медицинской микробиологии.
В одной из публикаций (Ж. микробиол., 1989, №10, с. 188) я выразил свое отношение к учебной литературе по медицинской микробиологии и сформулировал правила, с использованием которых, по моему мнению, ее следует готовить. Главный постулат очевиден — надо знать, кого и чему мы хотим научить. Есть академическая сторона дела и лабораторная служба. Врачи, по-видимому, больше заинтересованы в первом, т.е. в понимании сущности предмета. О технических деталях надо знать ровно столько, чтобы с уважением относиться к тем, кто создает и реализует блестящие методы микробиологического анализа.
Ясно и то, что книги из круга начального чтения должны отличаться от справочников. Их задача — учить не формальному, а аналитическому запоминанию, т.е. запоминанию через размышление. Основой фактологии должна быть концептуальность, философия предмета. Для этого материал обязан быть читабельным, в идеале приближаясь к хрестоматии. Чтение должно возбуждать интерес и стремление шире охватить предмет, нацеливая на специальные руководства, но не заменяя их, ибо «учение без размышления бесполезно» (В.Г. Белинский).
В книге сделан акцент на патогенетически значимые признаки возбудителей в процессах, которые они запускают, вынуждая хозяина к сопротивлению. Это сделано потому, что автор адресует свой труд врачам (настоящим и будущим), полагая, что стержнем медицинской микробиологии должно быть понимание механизмов инфекционного процесса. Свойства микробов, не имеющие прямого отношения к патогенезу, по возможности опущены. При желании нетрудно найти литературу, где об этом рассказано достаточно подробно.
Книга написана в форме очерков, ибо представленный материал не исчерпывает всех сюжетов инфекционной патологии, а его изложение не отвечает формальной дидактике. Она рассчитана на любознательного читателя, обладающего элементарными знаниями в области микробиологии, иммунологии, биохимии.
Автор глубоко признателен Нидерландскому фонду научных исследований, научно-производственной организации «Диагностические системы» (Н. Новгород) и факультету обучения иностранных студентов Нижегородской медицинской академии, благодаря финансовой поддержке которых он имел возможность следить за научной периодикой и монографическими изданиями по микробиологии и иммунологии.
Основная литература, использованная для иллюстраций и таблиц
- Авакян А.А., Кац Л.Н., Павлова И.Б. Атлас анатомии бактерий, патогенных для человека и животных. М.: Медицина, 1972.
- Salyers A.A., Whitt D.D. Bacterial pathogenesis. a molecular approach. 2nd ed. asm press. Washington, D.C., 2002.
- Topley & Wilson’s principles of bacteriology, virology and immunity (A.H. Linton & H.M. Dick, eds.). 8th ed. E. Arnold, 1990.
- Basic and clinical immunology (D.P. Stites , A.I. Terr, eds). 7th ed. Appleton & Lange, 1991.
- Medical microbiology (C.A. Mims et al., eds.). Mosby, 1993.
- Taussig M.J. Processes in pathology and microbiology. 2nd ed. Blackwell Sci. Publ., 1984.
- Burrow’s textbook of microbiology (B.A. Freeman, ed.). 22nd ed. W.B. Saunders Company, 1985.
- Mechanisms of microbial disease (M. Schaechter, G. Medoff, B.I. Eisenstein, eds.). 3rd ed. Williams & Wilkins, 1999.
- Koehler W., Mochman H. Grundris der medizinischer microbiologie. Veb Gustav Verlag Jena, 1975.
- Smith A.L. Principles of microbiology. 10th ed. Times Mirror/Mosby, 1995.
- Olds R.J. A colour atlas of microbiology. Wolfe Med. Publ. LTD, 1986.
- Color atlas and textbook of diagnostic microbiology (E. Koneman et al., eds.). 4th ed. J.B. Lippincott Co., 1992.
- Forbes C.D., Jackson W.F. A colour atlas and text of clinical medicine. Wolfe Med. Publ., 1993.
- Bergey’s manual of systematic bacteriology. 9th ed. Williams & Wilkins, 1984, Vol. 1.
- Internal medicine (J.H. Stein, ed.). 4th ed. Mosby, 1994.
- Clinical virology (D.D. Richman, R.J. Whitley, F.G. Hayden, eds.). 2nd ed. ASM Press, Washington D.C., 2002.
- Grigoriu D., Delacretaz J., Borelli D. Medical Mycology. H. Huber Publishers Toronto & Lewiston, N.Y.—Bern-Stuttgart, 1987.
- Manual of clinical microbilogy (P.R. Murray et al., eds.). 7th ed. ASM Press, Washington D.C., 1999.
- Madigan M.T., Martinko J.M., Parker J. Brock biology of microorganisms. 8th ed. Prentice Hall Intertionall, Inc., 1997.