
2006 год
Возбудитель дифтерии
- Клебс — Леффлер — Ру — Беринг.
- Биовары—патовары.
- Колонизация и интоксикация.
- Генетика и механизм действия дифтерийного токсина.
- Безвредные двойники и подражатели.
- Классика вакцинологии.
Дифтерия — острое инфекционное заболевание, символом которого является некротический фарингит с тяжелой интоксикацией, развивающейся через 2—10 дней после заражения. Деструкция эпителия, распространяющаяся на сосуды субэпителиальной ткани, ведет к массивной экссудации плазмы и выходу форменных элементов крови. Содержащийся в плазме фибриноген при контакте с тромбопластином некротизированного эпителия превращается в фибрин, который образует на слизистой оболочке пленку, прочно спаянную с подлежащей тканью. Такой тип воспаления называется дифтеритическим (греч. diphthera — кожа, пленка). Поражение глотки — самая распространенная форма заболевания, и большинство других его проявлений можно рассматривать как осложнения дифтерии зева. Процесс нередко захватывает низлежащие дыхательные пути, переходя в дифтерию гортани, трахеи, бронхов. Иногда инфекция распространяется вверх, поражая слизистую оболочку носа и среднего уха. Самостоятельными формами (если иметь в виду входные ворота инфекции) являются дифтерия половых органов (у девочек) и ран. Они встречаются редко и на фоне почти полного искоренения дифтерии в развитых странах практически забыты.
Самое страшное осложнение дифтерии — круп (шотл. croak — карканье; имитационный термин, подражающий хриплому, тяжелому дыханию больного). Это асфиксия, возникающая при закупорке респираторного тракта фибринозным экссудатом, который трудно откашливается и может полностью заблокировать дыхание. Если процесс распространяется на бронхи, то даже трахеотомия не всегда спасает больного. Отсюда понятны прежние названия дифтерии — смертельная язва глотки, удушающая болезнь, которыми пользовались еще врачи древности. В самостоятельную нозологическую единицу заболевание выделено в начале ХIХ в. французским врачом П. Бретонно и его учеником А. Труссо.
Наиболее тяжелый вариант общей интоксикации — гипертоксическая форма дифтерии. В этом случае на первый план выступает поражение миокарда, надпочечников, почек, центральной и периферической нервной системы. Смерть наступает при быстро нарастающем ослаблении сердечной деятельности. Но обычно симптомы развиваются медленнее, в течение 10—14 сут. У погибших определяются глубокие дистрофические изменения в жизненно важных органах — следствие отравления дифтерийным токсином, поступающим из очага инфекции.
В естественных условиях дифтерией болеет только человек (он же служит единственным резервуаром инфекции), хотя на клеточном (точнее субклеточном) уровне все эукариотические системы чувствительны к дифтерийному токсину.
Поиски возбудителя (Corynebacterium diphtheria) начались в лабораториях, руководимых Р.Кохом. В 1883 г. Э.Клебс обнаружил его в пленках, взятых из зева больных дифтерией, а спустя год он был выделен в чистой культуре Ф. Леффлером. Однако окончательное утверждение палочки Клебса—Леффлера в роли возбудителя дифтерии задержалось на несколько лет. Для убеждения скептиков потребовалось два эпохальных открытия. Сначала сотрудники Л.Пастера Э.Ру и А.Иерсен доказали токсичность фильтрата бульонных культур бактерий, изолированных Ф.Леффлером, а в 1888 г. Э. Беринг и С. Китазато (сотрудники Р. Коха) экспериментально обосновали способность сывороток животных, получивших сублетальные дозы дифтерийного и столбнячного токсинов, предупреждать гибель от введения тех же токсинов. Было сформулировано понятие об антитоксине, что явилось первым доказательством образования антител против чужеродных белков. В рождественскую ночь 1891 г. в одной из берлинских клиник П. Эрлих впервые ввел антитоксическую сыворотку ребенку, погибавшему от дифтерии. Блестящий результат этого отчаянного эксперимента обеспечил быстрое внедрение метода пассивной иммунизации в лечение дифтерии. До сих пор антитоксическая сыворотка служит единственным спасением от гипертоксических вариантов и местных осложнений дифтерии.
Эти открытия не только утвердили этиологическую значимость бактерий Клебса—Леффлера, но заставили по-новому взглянуть на патогенез инфекционных заболеваний, механизмы выздоровления и возможности их искусственного (иммунологического) усиления. Дифтерия занимает почетное место в истории бактериологии, являясь классическим примером инфекции с почти идеальным мономолекулярным механизмом патогенеза, абсолютно точными представлениями о генетических основах вирулентности возбудителя, способах специфической профилактики и терапии.
Родовое название возбудителя дифтерии, Сorynebacterium, отражает один из морфологических признаков этой группы бактерий: они имеют палочковидную форму с утолщениями на одном или обоих концах (греч. koryne — булава, дубинка). Клетки окрашиваются неравномерно из-за включений (зерен) волютина. Последние красятся метахроматически метиленовой синькой (желтая окраска на фоне голубого цвета клеток), на чем основаны специальные методы обнаружения дифтерийной палочки. С диагностической целью препараты обрабатывают одним из способов, дифференцирующих включения волютина (обычно по Леффлеру, слабо щелочным раствором метиленовой синьки). Коринебактерии грамположительны, но при окраске по Граму выявить зерна волютина (их называют зернами Бабеша—Эрнста) не удается.
В мазках дифтерийные палочки располагаются достаточно характерно: картину сравнивают с растопыренными пальцами, китайскими иероглифами, частоколом, латинскими буквами L, V, Y (рис. 1). Могут возникать ветвящиеся формы (особенно в старых культурах) — признак, по которому коринебактерии вместе с микобактериями некогда причисляли к актиномицетам. В целом складывается впечатление полиморфизма, который усиливается из-за неодинакового размера клеток (их длина колеблется от 1 до 8 мкм), а также из-за того, что в дифтеритических пленках и чистых культурах кроме одиночных клеток встречаются скопления бактерий, напоминающие войлок.

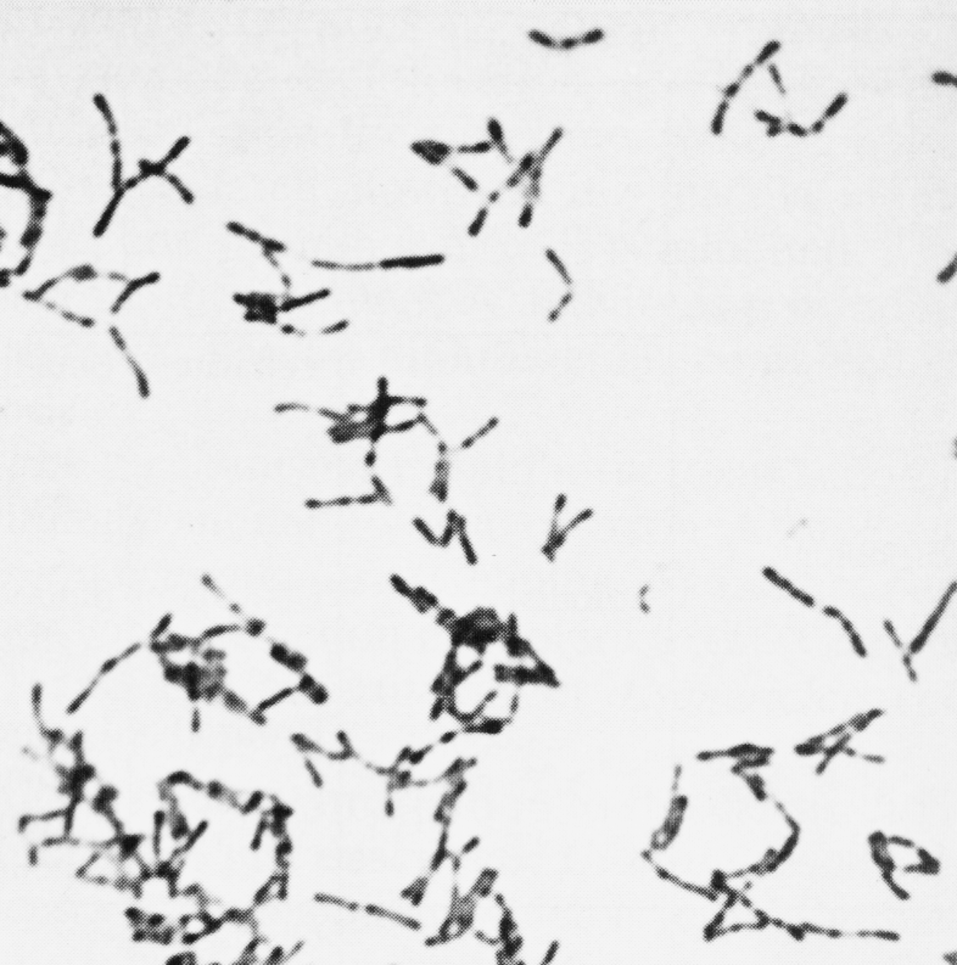
Неоднородность дифтерийной палочки находит отражение и в культуральных свойствах. По характеру роста на плотных питательных средах различают три биотипа (биовара) C. diphtheriae: gravis, intermedius и mitis. Колонии типа gravis формально напоминают R-форму бактерий (радиальная исчерченность, неровный край, без гемолиза). Колонии типа mitis больше похожи на S-форму (гладкие, выпуклые, с ровным краем, более мелкие, окружены зоной гемолиза). Тип intermedius занимает промежуточное положение, но по размеру колоний значительно уступает gravis и mitis (рис. 2). Биотипы дифтерийной палочки различаются и по ферментативной активности, причем gravis превосходит другие варианты. Немало усилий потрачено на то, чтобы связать различные типы дифтерийных бактерий с вирулентностью, превратив их из биоваров в патовары, т.е. в разновидности (варианты) с неодинаковой патогенностью. Этого сделать не удалось. Между культурально-биохимическими особенностями возбудителя и тяжестью заболевания нет четкой зависимости, и эти многолетние усилия представляют больше исторический интерес, отражая искренние (а порой и тенденциозные) заблуждения бактериологов, оставивших о себе память в ошибочных терминах — gravis (лат. тяжелый), mitis (легкий), intermedius (промежуточный).
Рис. 2. Колонии C. diphtheriae на кровяном теллуритовом агаре (48 ч, 37°С): а — тип gravis, б — тип mitis, в — тип intermedius


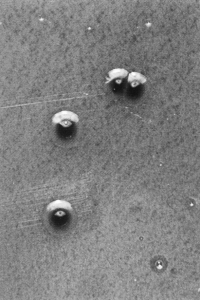
Подобно всем коринебактериям (их около 20 видов), дифтерийная палочка непритязательна к питательным средам, но ее предпочитают культивировать на средах с кровью или сывороткой, так как в первых генерациях (т.е. при выделении от больного) она растет скудно и лишь при повторных посевах привыкает к новому режиму. Кстати, успех Леффлера во многом предопределило использование среды, обогащенной сывороткой крови. Не создают проблем и условия культивирования. Дифтерийная палочка — факультативный анаэроб и отлично развивается в обычной атмосфере при 37°С. Культуры хорошо сохраняются: на свернутой сыворотке, защищенной от высыхания и света, возбудитель остается вирулентным более года. Словом, работа с дифтерийной палочкой не создает больших хлопот, тем более, что разработаны и производятся специальные коммерческие среды для выделения и идентификации возбудителя. Чаще пользуются кровяными средами с теллуритом, на которых колонии дифтерийной палочки легко дифференцировать от сопутствующей микрофлоры, тем более, что рост грамотрицательных бактерий в подобных условиях подавляется (см. рис. 2). Остальное дополняет микроскопия, изучение ферментативной активности и (главное!) способности к токсинообразованию.
Дифтерия принадлежит к немногим инфекциям, патогенетическую сущность которых определяет мономолекулярная интоксикация. Своевременное введение антител, нейтрализующих токсин (их вводят в виде антитоксической сыворотки или антитоксического иммуноглобулина), обрывает симптомы заболевания, а в опытах на животных предупреждает развитие местных поражений и смертельной интоксикации. Размножение бактерий почти всегда ограничивается слизистыми оболочками, не выходя за пределы эпителиального барьера. Однако токсин, секретируемый возбудителем, преодолевает этот рубеж и через регионарный лимфоидный барьер проникает в кровяное русло, поражая органы, которые наиболее интенсивно снабжаются кровью и содержат максимальное число рецепторов к токсину (см. ниже).
Дифтерийный токсин убивает клетки эпителия, подавляет фагоцитоз, а повреждая эндотелиоциты и стенку сосудов, вызывает обильную экссудацию. Экссудат свертывается, плотно спаиваясь с подлежащей тканью. Образуется толстая плотная пленка — псевдомембрана, содержащая сгустки фибрина, некротизированные ткани и бактерии. Возбудитель получает здесь великолепные условия для размножения и продукции токсина. Дифтерийная палочка продуцирует ферменты (нейраминидазу и N-ацетилнейраминиатлиазу), которые, действуя на эстафетной основе, обеспечивают бактерии энергетическим сырьем: нейраминидаза отщепляет N-ацетилнейраминовую кислоту от гликопротеинов слизи и поверхности клеток, а лиаза расщепляет ее на пируват и N-ацетилманнозамин. Пируват служит готовым источником энергии, стимулируя рост коринебактерий.
Не обладая капсулой, возбудитель находит внутри дифтеритической пленки отличную защиту от эффекторов иммунитета. Палочка дифтерии не образует спор, но в пленках не погибает при 98°С в течение часа, хотя чистая культура даже при 60°С гибнет через несколько минут. В высохшей пленке дифтерийная палочка сохраняет жизнеспособность около месяца, тогда как в чистом виде (например, на детских игрушках) выживает лишь несколько дней.
Выделив впервые культуру дифтерийной палочки, Ф. Леффлер не скрывал опасений, что его открытие может быть ложным. Причиной этих сомнений были противоречия с имевшими тогда непререкаемый авторитет постулатами Р. Коха, согласно которым микроб мог быть признан возбудителем болезни при трех обязательных условиях:
- он должен выделяться от всех больных данным заболеванием;
- обязан отсутствовать у здоровых и больных другими заболеваниями;
- основные симптомы должны быть воспроизведены при заражении чистой культурой экспериментальных животных.
Сомнения возникли прежде всего по второму пункту, так как бактерии, похожие на дифтерийную палочку (а иногда не отличимые от нее), выделялись из зева здоровых людей и недифтерийных больных. В этом быстро разобрались. Было установлено, что коринебактерии (они получили это название в 1896 г.) достаточно разнородны, и кроме истинной дифтерийной палочки человек инфицируется и другими представителями этой группы микроорганизмов. Их собирательно именуют ложнодифтерийными палочками, или дифтероидами. Есть четкие биохимические (ферментативные) и менее надежные (но все-таки значимые) морфологические и культуральные признаки, по которым истинные дифтерийные палочки отличаются от «псевдовидов».
Но оставалась другая загвоздка: путаницу вносили штаммы, которые по классическим маркерам невозможно отдифференцировать от патогенетически значимых изолятов дифтерийной палочки. Проблема решилась с открытием дифтерийного токсина. Оказалось, что единственное отличие безвредных двойников от вирулентных штаммов состоит в том, что они не продуцируют токсин, т.е. являются нетоксигенными. Нетоксигенные штаммы не вызывают дифтерии, хотя и способны персистировать в респираторном тракте человека. По сути, это факультативные представители нормальной микрофлоры, а бессимптомность инфекции доказывает, что токсинообразование — не обязательный (или, по крайней мере, не единственный) фактор колонизации.
Загадка токсигенных и нетоксигенных двойников дифтерийной палочки прояснилась в 1951 г. Оказалось, что токсигенные штаммы заражены умеренными фагами (b- и w-фаги), которые вносят в клетку tox-ген, ответственный за синтез токсина. Штаммы, не инфицированные лизогенными фагами или потерявшие их, лишены токсигенности. Бактериальная клетка сохраняет определенный контроль над токсинообразованием, продуцируя особый белок, который в присутствии ионов железа (Fe2-) подавляет транскрипцию tox-гена. Если содержание железа в среде падает ниже определенного уровня, образование токсина усиливается. Этим объясняется повышенный синтез токсина на поздних стадиях роста бактериальной культуры, когда основная часть неорганического железа израсходована. Сходная ситуация возникает, по-видимому, и в очагах дифтерийного поражения (в псевдомембранах), способствуя интоксикации.
Наконец, сомнениям Леффлера способствовало и то, что «классическая» дифтерийная палочка иногда обнаруживается в зеве здоровых людей. Но и это противоречие вскоре отпало: носители токсигенных штаммов защищены от токсина антитоксическими антителами. Поэтому они не болеют, но служат наиболее значимым резервуаром дифтерийной инфекции. Число носителей в разные годы и в разных регионах неодинаково (это во многом зависит от масштабов активной иммунизации) и, по сильно усредненным данным, составляет 0,2—0,5%. С практической точки зрения различают транзиторное носительство (1—7 дней), кратковременное (7—15 дней), средней продолжительности (15—30 дней) и затяжное (более месяца).
Дифтерийный токсин относится к токсинам, которые проявляют ферментативную активность внутри поражаемых клеток. Он принадлежит к категории АДФ-рибозил-трансфераз, т.е. ферментов, переносящих АДФ-рибозу (она отщепляется от НАД) на акцепторные белки-мишени, блокируя их биологическую (ферментативную) активность (см. «Болезнетворность бактерий»). Дифтерийный токсин АДФ-рибозилирует фактор элонгации-2 (EF-2), который необходим для построения пептидных цепей на рибосомах эукариотических клеток. Его блокада подавляет синтез белка и вызывает гибель клеток (рис. 3). Прокариотические клетки используют другой фактор элонгации и поэтому не чувствительны к дифтерийному токсину.


Подобно всем бактериальным токсинам, действующим внутриклеточно, дифтерийный токсин является бинарной молекулой, т.е. состоит из двух фрагментов — А и В, выполняющих соответственно ферментативную (расщепление НАД, АДФ-рибозилирование) и рецепторную функции. Фрагмент В связывается с рецепторами клеток (гепаринсвязывающим эпидермальным фактором роста), способствуя внутриклеточному транспорту А-компонента. Этому сопутствует расщепление молекулы токсина мембранными протеазами и высвобождение активированного А-фрагмента. Полагают, что высокая чувствительность к дифтерийному токсину миоцитов сердца и нервных клеток объясняется повышенным содержанием в них гепаринсвязывающего эпидермального ростового фактора.
Определение токсигенности изолированной культуры венчает микробиологический анализ при подозрении на дифтерию: лишь выделение токсигенных культур имеет диагностическое значение. Дифтерийная палочка активно продуцирует токсин, поэтому для его обнаружения вполне достаточно не очень чувствительного, но весьма демонстративного метода двойной (встречной) иммунодиффузии в геле, который по автору называется реакцией Оухтерлони (рис. 4).

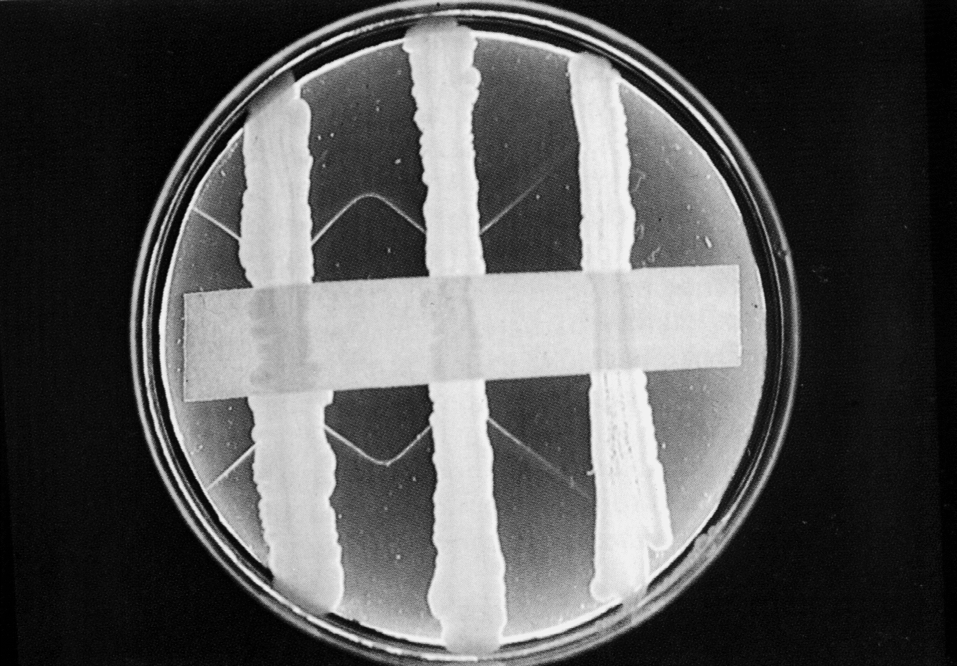
Представления о дифтерии как о специфической интоксикации сыграли решающую роль в борьбе с этой инфекцией. Выздоровление связано с образованием антител, нейтрализующих токсин. Этому можно содействовать, вводя больному готовые антитела в виде антитоксической сыворотки или ее иммуноглобулиновой фракции. Антитоксическую сыворотку получают от гипериммунизированных лошадей, так что следует считаться с возможностью аллергических осложнений на введение чужеродного (гетерологичного) белка. Они бывают разной тяжести и наблюдаются примерно в 10% случаев. Но это терапия выбора и начинать ее следует как можно раньше, так как, нейтрализуя свободный токсин, антитела бессильны против токсина, связавшегося с клетками, а тем более уже проникшего в них. Это означает, что сигналом к иммунотерапии должен быть клинический диагноз дифтерии. Выделение токсигенной культуры служит хоть и важным, но все-таки запоздалым подтверждением. Антибиотики приносят пользу, но не заменяют антитоксина.
Блестящим развитием идеи о дифтерийной интоксикации явилась разработка подходов к искусственному созданию активного иммунитета против дифтерии. В 1920-х гг. французским ученым Г. Рамоном разработан способ обезвреживания (детоксикации) дифтерийного токсина с сохранением его иммуногенных свойств. Такой препарат (он называется анатоксином) получают путем обработки токсина 0,3% раствором формальдегида, который сшивает остатки лизина и тирозина, образуя метиленовые мостики. Анатоксин не связывается с клеточными рецепторами, не обладает АДФ-рибозил-трансферазной активностью и не расщепляется на А—В-фрагменты. Этим объясняется его полная безвредность. Глобальное применение анатоксина для вакцинации детей (его обычно применяют в виде преципитата на гидроокиси алюминия — адсорбированный анатоксин) привело к резкому сокращению дифтерии. Многочисленными исследованиями доказано, что восприимчивость к дифтерии зависит от уровня антитоксического иммунитета, и лишь нарушения в вакцинной практике срывают программу ликвидации этой инфекции. Это, в частности, привело к недавнему росту заболеваемости дифтерией на территории бывшего Советского Союза. Так, в 1994 г. было зафиксировано почти 50 000 случаев дифтерии, из которых 1750 закончились смертельным исходом. Болели преимущественно взрослые и дети старшего возраста, утратившие иммунитет и не восполнившие его дефицит путем своевременной ревакцинации.
Антитоксические антитела с опережением нейтрализуют токсин, образующийся во входных воротах инфекции. Это исключает повреждение тканей, предупреждая образование фибринозных пленок, которые стабилизируют инфекцию, способствуя ее патогенетически значимой эволюции. Без такой опоры дифтерийная палочка обычно надолго не задерживается на слизистых оболочках. Впрочем, как уже говорилось, примеры бессимптомного носительства токсигенных штаммов дифтерийной палочки на фоне протективных титров антитоксических антител, а также присутствие в зеве ее нетоксигенных вариантов говорят о том, что токсин не является обязательным фактором колонизации. Совершенно очевидно, что подобно другим коринебактериям дифтерийная палочка располагает поверхностными структурами, которые обеспечивают ее адгезию на эпителиоцитах. Их природа не известна, но это рождает естественное стремление расширить тактику превентивной борьбы против дифтерии. Главные идеи связаны с предупреждением колонизации слизистых оболочек и эпидемиологически значимого носительства возбудителя. Надежды возлагаются на вакцинные препараты из компонентов самих бактерий, т.е. на создание антибактериального иммунитета, который смог бы усилить элиминирующие потенции антитоксических антител. Хочется верить, что эта логика принесет плоды, но она должна базироваться на грамотных представлениях о факторах микробной колонизации и не повторять тщетных попыток прошлого. У человечества уже есть «синица в руках» — анатоксин, который отлично зарекомендовал себя в борьбе с дифтерией. Не исключено, что «журавль в небе» не понадобится, если организованно поддерживать классический опыт анатоксиновой иммунопрофилактики.
Микобактерии туберкулеза
По опустошительному своему действию туберкулез занимает первое место среди других болезней.
Энциклопедический словарь Брокгауза—Эфрона, 1902
- Экологические разновидности микобактерий.
- Клеточная стенка: микробиологическое и патогенетическое своеобразие туберкулезной палочки.
- «Болезнь макрофагов».
- Туберкулезная гранулема: эволюция неспецифических и специфических механизмов.
- Аллергия и иммунитет.
- Болезнетворность: прямые и опосредованные эффекты.
- Персистенция, реактивация и патогенетические варианты туберкулеза.
- Специфическая диагностика: возможности, ограничения, перспективы.
- Вакцинопрофилактика: иллюзия безопасности.
- Этиотропная терапия: классика и тревоги будущего.
Название болезни «туберкулез» происходит от характерного патоморфологического признака — специфической гранулемы, или туберкула. Это очаг хронического, иммунологически зависимого воспаления, которое в типичном виде выглядит как небольшой бугорок (лат. tuberculum), внешне и гистологически отличающийся от гранулем иного происхождения. Слово «туберкул» впервые употребил в XVI в. Ф.Сильвий при описании поражений легких у людей, умерших от чахотки (греч. phtisis — истощение; отсюда — фтизиатрия, фтизиатр). В качестве самостоятельного понятия «туберкулез» стали использовать в XIX в. после того, как Ж.Бейль и Р.Лаэннек доказали, что образование туберкулов — непременный спутник болезни.
Туберкулез известен с глубокой древности и почти наверняка причинил человечеству больше страданий, чем любая другая инфекция. Несмотря на блестящие достижения этиотропной терапии, он и сегодня остается тревожной проблемой. Прогноз ВОЗ в 1960 г. о том, что туберкулез в ближайшее время будет ликвидирован как широко распространенное заболевание, не состоялся, и уже в 1993 г. было заявлено, что туберкулезная инфекция выходит из-под контроля. В современных публикациях туберкулез часто упоминается среди так называемых возрождающихся и вновь возникающих (англ. emerging/reemerging) инфекций. Ежегодно на Земном шаре от туберкулеза умирает более 3 млн. человек, и вряд ли следует ожидать быстрого улучшения ситуации.
Догадки о заразности туберкулеза восходят к далекому прошлому, но лишь в 1868 г. Ж. Вильмену удалось воспроизвести заболевание у животных при заражении мокротой от больных туберкулезом. Удивительно, но эти исследования (как и более поздние работы Ю.Конгейма) не убедили современников. Туберкулез продолжали связывать с самыми невероятными «капризами природы», и лишь работы Роберта Коха утвердили его инфекционное начало.
В марте 1882 г. на заседании Берлинского физиологического общества Кох сообщил о своих исследованиях по этиологии туберкулеза. В мокроте туберкулезных больных он обнаружил палочки, которые по тинкториальным свойствам и росту на питательных средах отличались от всех известных тогда бактерий. Заражение чистыми культурами вызывало туберкулезный процесс у животных. В докладе было немало и других сведений о происхождении одной из самых страшных болезней человека. Нельзя не поражаться огромной творческой силе, которая потребовалась Коху, чтобы выполнить поставленные перед собой задачи. Дело в том, что туберкулезная палочка не выявляется при обычной окраске, растет во много раз медленнее других бактерий, а ее выделение в чистой культуре представляет трудности даже для современных бактериологов. Исследования по туберкулезу явились главным основанием для присуждения Р. Коху одной из первых Нобелевских премий по медицине.
Таксономия и разновидности
Туберкулез человека вызывают два вида микобактерий — Mycobacterium tuberculosis и Mycobacterium bovis (микобактерии бычьего типа). Их нетрудно различить по ряду признаков, но этого часто не требуется, так как современный туберкулез обычно связан с М. tuberculosis.
Если быть точнее, следует говорить о так называемом туберкулезном комплексе, или группе М. tuberculosis. Кроме М. tuberculosis и М. bovis сюда входят еще два вида микобактерий — М. africanum и М.microti. Из них лишь М.africanum изредка вызывает туберкулез у человека, причем не только среди жителей Африки (как следует из видового эпитета), но и в других регионах. Памятуя о микробиологической неоднозначности туберкулеза, мы тем не менее будем ориентироваться на главный вид, М. tuberculosis, с которым связано более 90% случаев туберкулеза, регистрируемых на Земном шаре. Кроме того, М. tuberculosis и М. bovis так похожи, что некоторые специалисты до сих пор считают их вариантами одного вида.
Заражение микобактериями бычьего типа происходит при употреблении молока больных коров, и ветеринарный контроль за животными в сочетании с пастеризацией молока практически исключают опасность для человека. Эти простые мероприятия, ставшие очевидными после признания М. bovis самостоятельным видом микобактерий, привели к резкому сокращению «коровьего» туберкулеза, став одним из блестящих достижений чисто санитарной эпидемиологии. Полной ликвидации М.bovis мешает его циркуляция среди многочисленных видов диких животных.
Родовое название Mycobacterium (грибовидные бактерии) предложено Леманом и Нойманом в 1896 г., обративших внимание на плесневидную пленку, которая образуется при росте туберкулезной палочки на поверхности жидких сред (аэробы!). Вскоре коллекция микобактерий была дополнена видами, изолированными из внешней среды. Оказалось, что именно свободно живущие сапрофиты составляют большинство этой своеобразной группы прокариот, а истинно паразитические микобактерии представлены всего несколькими видами, патогенными для человека и животных. Впрочем, известную опасность представляют и некоторые микобактерии-сапрофиты (рис. 1). Они вызывают оппортунистические инфекции (микобактериозы) у иммунокомпрометированных лиц, а при заражении в раннем детстве могут извращать реактивность организма к возбудителям туберкулеза.

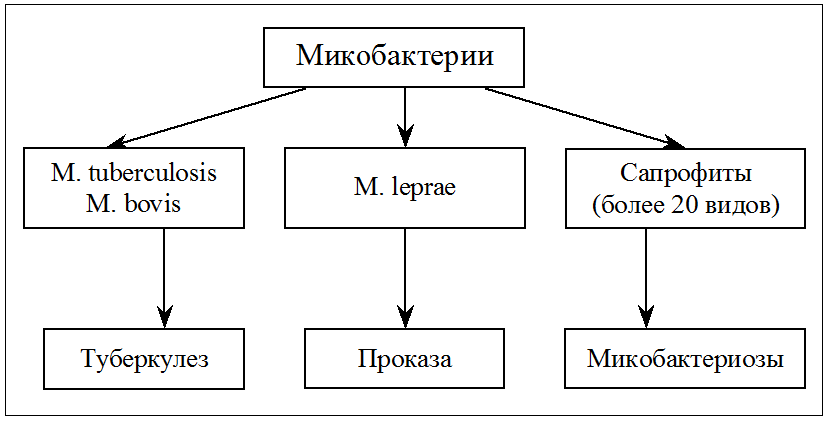
Известно более 60 видов микобактерий. Они объединены в род Mycobacterium семейства Mycobacteriaceae, которое относится к порядку Actinomycetales. В этом есть логика. Подобно актиномицетам, микобактерии образуют ветвящиеся клетки (особенно in vitro). Но сходство имеет и более глубокие корни, по крайней мере с соседями по порядку — нокардиями и коринебактериями. И те и другие содержат липиды, которые напоминают миколовые кислоты, хотя и уступают микобактериям по сложности строения этих уникальных компонентов клеточной стенки.
Морфология, тинкториальные свойства
Микобактерии туберкулеза представляют собой тонкие, прямые или слегка изогнутые палочки, длиной 1—4 мкм и около 0,3 мкм в ширину (рис. 2). Они неподвижны, не образуют спор и капсулы, если не считать микозидной оболочки, которую иногда сравнивают с микрокапсулой. Они плохо окрашиваются по Граму, но, восприняв окраску, не обесцвечиваются этанолом, поэтому их считают грамположительными, хотя правильнее не относить ни к тем, ни к другим. С трудностями окраски туберкулезной палочки впервые столкнулся Кох, сумевший разглядеть их в мазках из мокроты лишь после 24—30-часовой (!) инкубации в щелочном растворе метиленовой синьки.

Избирательная окраска основана на так называемой кислотоустойчивости микобактерий и включает два основных этапа. Сначала повышают проницаемость клеточной стенки для насыщенного раствора красителя (расплавление восковидного слоя путем прогревания или протравливания детергенами), а затем окрашенный препарат промывают разведенными кислотами. Обесцвечивая большинство бактерий, это сохраняет окраску микобактериальных клеток. По классической методике Циля—Нильсена на фиксированный мазок наливают карболовый фуксин, нагревают до отхождения паров, промывают закисленным этанолом (3% НСl в 85% растворе этанола) и докрашивают метиленовой синькой. Микобактерии удерживают фуксин и выглядят как красные палочки на голубом фоне; остальные бактерии теряют фуксин и окрашиваются в синий цвет. Этанол здесь не обязателен, но делает фон более чистым. Устойчивость к обесцвечиванию (она связана с образованием прочных комплексов между красителем и миколовыми кислотами клеточной стенки) проявляется не только в отношении кислот, но также щелочей и спиртов. Поэтому с равной справедливостью микобактерии можно называть также щелоче- и спиртоустойчивыми.
Морфологические и тинкториальные признаки микобактерий имеют много исключений, породивших немало споров о природе возбудителя. Туберкулезная палочка может терять кислотоустойчивость и хорошо окрашиваться по Граму, превращаться в филаментозные и даже мицелиоподобные формы, распадающиеся на палочки и кокки (гранулы/зерна Муха). Некоторые из них настолько малы, что проходят через бактериальные фильтры (фильтрующиеся формы). Теперь ясно: речь идет о фенотипической изменчивости, которая отражает экологическую пластичность возбудителя и его способность выживать в неблагоприятных условиях. Отсюда не удивительно, что плеоморфизм туберкулезной палочки сильнее проявляется в инвитровых культурах, т.е. в неестественной среде обитания.
Культуральные особенности
Туберкулезные микобактерии — строгие аэробы и мезофилы, т.е. растут в диапазоне 30—42° С, лучше всего при 37°С. Размножение происходит очень медленно: период генерации составляет 14—16 ч (типичные бактерии делятся каждые 15 мин). Поэтому для получения обильного роста требуется не менее 4—6 нед, хотя миниатюрные колонии могут появиться через 7—10 дней. Туберкулезная палочка принадлежит к числу наиболее вяло реплицирующихся микобактерий. Большинство сапрофитических видов размножаются быстрее, их рост заметно отстает от других прокариот и хорошо заметен не ранее чем через 5—7 дней. Одной из причин отсроченного размножения микобактерий является высокая гидрофобность, связанная с обилием липидов в клеточной стенке. Это затрудняет поступление в бактерии питательных веществ, снижая их метаболическую активность.
Выделение первичных культур (т.е. непосредственно от больного) проводят на специальных средах, самые сложные из которых содержат яйца, картофельную муку и глицерин. Для подавления сопутствующей микрофлоры добавляют малахитовый зеленый или генцианвиолет. При субкультивировании туберкулезная палочка становится менее прихотливой и растет на обычных средах с добавкой глицерина.
В жидких средах рост происходит на поверхности (аэроб!). Нежная сухая пленка со временем утолщается, становится бугристо-морщинистой и обретает желтоватый оттенок, часто сравниваемый с цветом слоновой кости. Бульон остается прозрачным и добиться диффузного роста удается только в присутствии детергентов, например твина-80. В микроколониях (они образуются на ранних сроках и заметны только под микроскопом) формируются структуры, напоминающие жгуты — признак, который связывают с так называемым корд-фактором М. tuberculosis (рис. 3).


Внутривидовые варианты
Морфотинкториальная изменчивость, о которой говорилось выше, весьма характерна для туберкулезной палочки, но это — фенотипические варианты, которые не переходят в генетически закрепленные биовары. Попытки выявить устойчивые серотипы М. tuberculosis и М. bovis не увенчались успехом, прежде всего из-за спонтанной агглютинации клеток, которая является следствием их высокой гидрофобности (см. ниже). Более удачным оказался опыт фаготипирования, но и он не закрепился в лабораторной практике.
В современных исследованиях много внимания уделяется дифференцировке (клонированию) микобактериальных штаммов по генетическим маркерам, прежде всего по особенностям хромосомного профиля ДНК. Генотипирование обычно проводится на основе вставочных генов — инсерционных последовательностей (англ. insertion sequences — IS). Они отличаются структурным полиморфизмом, позволяя классифицировать штаммы по степени генетического родства. Чаще используется анализ последовательности IS6110. Применяются и другие, дополнительные методы, основанные на особенностях генетического аппарата М. tuberculosis. К их числу относится сполиготипирование (от англ. spacer oligotyping), сравнение по числу прямых повторов и полиморфной G-С-обогащенной повторяющейся последовательности.
Полная идентичность возможна лишь внутри одного клона бактерий. Отсюда сравнение с отпечатками пальцев — англ. DNA fingerprinting. Каждое очередное поколение (т.е. каждый новый клон) несет хотя бы небольшие генетические различия. Накапливаясь со временем, они ведут к формированию клоногрупп, объединяемых в семейства. Тенденцию к распространению имеют клональные кластеры, еще не достигшие статуса группы. Наиболее изученным и распространенным (в том числе в России) является W-Beijing-семейство штаммов микобактерий туберкулеза. Оно включает более десятка клоногрупп — генетических ответвлений, эволюционировавших по IS6110 в различных географических зонах. Факторы, содействующие селекции штаммов W-Beijing, не известны. Возможно, это связано с повышенной контагиозностью, устойчивостью во внешней среде, резистентностью к антибиотикам. Именно первый W-штамм (акроним выбран произвольно), изолированный в 1990 г. в США, открыл тревожную эру лекарственной полирезистентности микобактерий туберкулеза. Есть мнение, что W-Beijing-штаммы более удачливы в преодолении барьера, создаваемого BCG-вакцинацией. Впрочем, каждая из перечисленных позиций встречает возражения.
Большую озабоченность вызывает эволюция лекарственной устойчивости на основе генетически закрепленных мутаций М. tuberculosis. Распространение резистентных биоваров может обезоружить в борьбе с туберкулезом или, по крайней мере, заметно снизить ее эффективность. Не случайно в системе лабораторий США, работающих с возбудителями особо опасных инфекций, организован центр, единственная задача которого — изучение штаммов туберкулезной палочки с множественной устойчивостью к антибиотикам.
Клеточная стенка
Разгадка своеобразия микобактерий связана с необычностью их поверхностных структур. В клеточной стенке, устроенной сложнее, чем у других бактерий, преобладают липиды (более 60% сухой массы), в том числе специфичные для микобактерий. Именно они определяют нестандартность тинкториальных, физиологических и экологических свойств туберкулезной палочки (см. таблицу). Разнообразие микобактериальных липидов, в которых с трудом ориентируются даже искушенные биохимики, заставляет пользоваться такими собирательными понятиями, как миколовые кислоты, микозиды, сульфолипиды, корд-факторы и пр.
| Признак | Причина |
| Тинкториальные свойства: | |
| неокрашиваемость обычными способами | Слабая проницаемость клеточной стенки |
| кислото-, щелоче-, спиртоустойчивость | Связывание красителей миколовыми кислотами |
| Медленное размножение | Низкая скорость внутриклеточной диффузии питательных веществ |
| Культуральные свойства (сухие, морщинистые колонии), спонтанная агглютинация бактериальных клеток | Гидрофобность клеток |
| Устойчивость во внешней среде | Защита от высыхания |
| Устойчивость к дезинфектантам | Медленное проникновение антисептиков в клетку |
| Взаимоотношения с макрофагами: | |
| поглощение | Гидрофобность клеток |
| внутриклеточное выживание | Блокада образования фаголизосом, нейтрализация антимикробных факторов, ускользание в цитоплазму (повреждение фагосомальных мембран) |
| Цитотоксичность | Повреждение митохондриальной мембраны |
| Особенности иммунитета, иммунопатогенез | Иммуноадъювантная активность, CD1-зависимое представление антигенов (гликолипидов) |
Большинство липидов представлено миколовыми кислотами и их производными — длинноцепочечными (60—90 углеродных атомов), разветвленными жирными кислотами. Нечто подобное есть только у коринебактерий и нокардий, но корино- и нокардиомиколовые кислоты гораздо короче — соответственно 28—40 и 40—60 углеродных атомов. Часть миколовых кислот ковалентно связана с пептидогликаном посредством арабиногалактана. При экстракции хлороформом его получают в виде фракции «воск D». Миколовые кислоты образуют подобие палисада, определяя восковидность структуры. Но миколовые кислоты не только фиксированы в каркасе клеточной стенки. Они присутствуют и в виде свободных гликолипидов-сульфолипидов (сульфатидов) и корд-фактора (точнее корд-факторов), едва ли не самого знаменитого из микобактериальных липидов. Это трегалоза (дисахарид глюкозы), эстерифицированная двумя молекулами миколовых кислот (димиколат трегалозы). По укоренившемуся представлению, корд-фактор определяет рост М. tuberculosis в виде серпантинных жгутов (англ. cord — жгут, веревка; см. рис. 3). Этому есть экспериментальные подтверждения, хотя непонятно, почему такие микроколонии не образуют другие микобактерии, содержащие сходные факторы. Из учебника в учебник переписывается положение о том, что корд-фактор — чуть ли не главный фактор вирулентности туберкулезной палочки. Однако для этого нет достаточных оснований. В клеточных культурах корд-фактор токсичен для биологических (особенно митохондриальных) мембран, но это зависит от вспомогательных факторов, которых могут быть лишены невирулентные штаммы М. tuberculosis и непатогенные микобактерии. На эту роль претендуют, в частности, сульфатиды (трегалоза, эстерифицированная сульфатной группой и четырьмя миколовыми кислотами), образующие мембранотропные цитотоксические комплексы с корд-фактором. Впрочем, туберкулезная палочка не утрачивает вирулентности и при почти полном отсутствии сульфатидов. Такого рода противоречия побуждают искать более сложные механизмы болезнетворности — прямые и опосредованные.
Уникальность и ключевая роль миколовых кислот в структурной организации и физиологии микобактерий делают их отличной мишенью для этиотропной терапии. Действительно, изониазид и этионамид, которые относятся к числу наиболее эффективных противотуберкулезных средств, блокируют синтез миколовых кислот М. tuberculosis. Удивительно, но они беспомощны против возбудителей микобактериозов, а этионамид к тому же слабо действует на М. bovis. Не отрицая других причин (например, неодинаковой способности к активирующей трансформации антибиотиков), можно думать об особенностях липидного синтеза у микобактерий. Этому соответствует разнообразие видовых спектров миколовых кислот, которое учитывается в таксономике.
Снаружи клеточная стенка окутана, подобно паутине, слоем гликолипидов. Их собирательно называют микозидами и иногда сравнивают с микрокапсулой. Микозиды структурно и функционально напоминают липополисахариды наружной мембраны грамотрицательных бактерий, но лишены их агрессивности. Тем не менее, они токсичны и, подобно корд-фактору и сульфатидам, вызывают образование гранулем.
Еще одним компонентом клеточной стенки микобактерий является липоарабиноманнан. Он заякорен на плазматической мембране, пронизывает клеточную стенку и выходит на ее поверхность. В этом отношении он похож на липотейхоевые кислоты грамположительных бактерий. Липоарабиноманнан представлен гетерогенной смесью высокомолекулярных липополисахаридов, углеводным компонентом которых служат разветвленные полимеры арабинозы и маннозы, а липидная часть состоит из диацилглицерольных производных пальмитиновой и туберкулостеариновой кислот. Особенности терминального фрагмента липоарабиноманнана (прежде всего его маннозные радикалы — кэпы) существенно влияют на взаимоотношения микобактерий с макрофагами, вмешиваясь в патогенез туберкулезного процесса.
Не следует забывать и о белках, которые функционируют в составе клеточной стенки, оживляя ее запутанную (для исследователей, но не для бактерий!) архитектонику. Часть из них входит в состав диагностического препарата «туберкулин» и претендует на роль антигенов для включения в вакцину. Особую известность получил протеиновый комплекс под названием «антиген 85». Он состоит из трех близкородственных белков, которые относятся к числу сильных индукторов иммунного ответа против M. tuberculosis. Данный комплекс выполняет функцию фермента, соединяющего миколовые кислоты с трегалозой. Это делает его мишенью для новых антибактериальных препаратов, которые в настоящее время проходят испытания.
Устойчивость во внешней среде
Туберкулезные палочки мало отличаются от других, не образующих споры бактерий по чувствительности к прогреванию, но в мокроте при кипячении погибают лишь через 5 мин. Характерна высокая устойчивость к высушиванию и химическим дезинфектантам, что скорее всего объясняется высоким содержанием липидов. Сухой жар (100°С) убивает их через 60 мин. В мокроте остаются жизнеспособными многие недели и даже месяцы. Контагиозность сохраняется 8—10 дней после высыхания и распыления мокроты в воздухе — именно столько туберкулезная палочка живет на рассеянном свету. Прямой солнечный свет служит хорошим дезинфектантом, убивая ее в течение часа. Фенол проникает в микобактерии медленно: для уничтожения возбудителя в мокроте требуется не менее 24 ч. Другие антисептики действуют тоже отсроченно, а некоторые практически не эффективны.
Факторы и механизмы туберкулезного процесса
Заражение. Чаще всего М. tuberculosis проникает в организм в составе мелкодисперсных аэрозолей. Для возбуждения конфликта с хозяином бактерии должны попасть в альвеолы. Это удается частицам диаметром 5—10 мкм; при более крупном размере они задерживаются в респираторном тракте и удаляются путем ретроградного мукоцилиарного транспорта. В альвеолах туберкулезная палочка поглощается резидентными (т.е. постоянно присутствующими здесь) макрофагами, взаимоотношения с которыми определяют дальнейшее развитие событий.
Взаимоотношения с макрофагами. Болезнетворность основана на способности микобактерий туберкулеза выживать и реплицироваться в макрофагах. Не случайно туберкулез относится к классическим внутримакрофагальным инфекциям. Склонность к внутримакрофагальной локализации наводит на противоречивые размышления. С одной стороны, это логично, если учесть 1) долгожительство мононуклеарных фагоцитов (это обеспечивает экологическую стабильность, необходимую медленно размножающимся микобактериям), 2) изоляцию от эффекторов гуморального иммунитета и 3) богатые пищевые ресурсы внутри клеток. В то же время нельзя не удивляться тому, что возбудитель выбирает биотоп, который из-за потенциальной опасности неприемлем для большинства микробов. Это одно из удивительных проявлений симбиоза — изощренного и во многом парадоксального.
Способы, благодаря которым внутримакрофагальные паразиты избегают уничтожения, связаны с ускользанием от гибельного воздействия фагоцитов. Главными условиями являются:
- незаметное (без активации макрофагов) проникновение в клетку;
- подавление образования фаголизосом или превращение их в зону, комфортную для бактерий;
- выход из фагосом в цитоплазму;
- устойчивость к антимикробным факторам или их инактивация;
- ослабление чувствительности макрофагов к активирующим сигналам Т-лимфоцитов;
- снижение антигенпредставляющей функции макрофагов и сопряженное с этим ослабление реакций цитотоксических Т-лимфоцитов, настроенных на уничтожение зараженных клеток.
В той или иной мере туберкулезная палочка использует каждый из этих механизмов.
Усредненная картина сводится к следующему. Туберкулезная палочка внедряется в макрофаги после связывания с поверхностью клеток. Главная роль принадлежит микозидам и липоарабиноманнану, которые напрямую воспринимаются клеточными рецепторами. Некоторые из таких рецепторов (например, рецепторы для маннозы) больше представлены на покоящихся (нестимулированных) клетках, а запускаемые через них сигналы не активируют (или слабо активируют) макрофаги. Поэтому фагоцитоз не сопровождается образованием таких мощных антимикробных факторов, как свободно-радикальные формы кислорода и оксид азота. Есть данные, что в большей степени это характерно для вирулентных штаммов М. tuberculosis, которые благодаря структурным особенностям липоарабиноманнана (наличие терминальных маннозных радикалов-кэпов) инициируют неагрессивный фагоцитоз. Авирулентные штаммы не столь коварны: они менее охотно взаимодействуют с маннозными рецепторами, теряя часть своего «инкогнито».
Выводы из модельных экспериментов (хотя и весьма изящных) нелегко проецировать на ситуации in vivo. В распознавание М. tuberculosis включаются и другие рецепторы макрофагов, в частности CD14, толльподобные рецепторы (TLR-1,2) и рецепторы для СЗ-фактора комплемента (CR1—CR3). В последнем случае, кроме прямых, имеют значение опосредованные реакции. Они возбуждаются производными СЗ-фактора, который фиксируется на поверхности бактерий. Этот механизм срабатывает без антител, т.е. через альтернативный механизм активации комплемента. Подспорьем для закрепления на клетках служит и высокая гидрофобность микобактерий, которая обеспечивает неспецифические (рецепторнезависимые) контакты с макрофагами.
Оказавшись внутри клеток, возбудитель приступает к очередным обезоруживающим акциям. Сначала туберкулезная палочка препятствует слиянию фагосом с лизосомами, где сконцентрирован основной запас антимикробных факторов. Этому способствуют защелачивание внутрифагосомальной среды аммонием, который продуцируется микобактериями, и миколовые кислоты, особенно сульфатиды, действующие благодаря высокому отрицательному заряду. Возможен более откровенный маневр: бактерии прорывают фагосомальную мембрану и выходят в цитоплазму, скрываясь здесь от биоцидных факторов. Скорее всего, это итог совместного действия сульфатидов и корд-фактора, образующих комплексы, токсичные для биомембран. В целом перегрузка микобактериальными липидами — нелегкое испытание для макрофага. Она чревата повреждением жизненно важных клеточных структур, которое может оказаться смертельным для клетки (цитолиз, апоптоз).
Но и внутри фаголизосом туберкулезные палочки не обречены на гибель. Благодаря мощной липидной оболочке, снискавшей им славу бронированных чудовищ, микобактерии малочувствительны к биоцидным началам фагоцитов. Их поверхностные гликолипиды (микозиды) сглаживают респираторный взрыв и инактивируют кислородные радикалы, аммоний защелачивает среду, блокируя активность лизосомальных ферментов, а сульфатиды нейтрализуют мембранотропные катионные пептиды. Кроме того, туберкулезная палочка продуцирует факторы с каталазной и пероксидазной активностями. Это повышает ее устойчивость к оксидантному стрессу.
Существенную роль в выживании туберкулезной палочки играют сидерофоры — бактериальные факторы, конкурирующие с хозяином за железо. Для микобактерий это особенно важно, так как, будучи аэробами, они должны активно синтезировать железосодержащие ферменты. Макрофаги (наряду с гепатоцитами) являются главным резервуаром железа, избыток которого резервируется в виде ферритина. Содержание железа в альвеолярных макрофагах в 100 раз выше, чем в моноцитах крови, что, безусловно, содействует их колонизации микобактериями туберкулеза. Последние обладают уникальным тандемом сидерофоров — экзохелином и микобактином. Экзохелины (пептиды) извлекают ионы железа из микроокружения и передают микобактинам (липиды клеточной стенки), которые обеспечивают накопление и транспорт железа внутрь бактерий.
Таким образом, для выживания внутри макрофагов туберкулезная палочка использует сложную тактику, и только совокупность различных приемов обеспечивает ее агрессивность. Этим вирулентные штаммы отличаются от авирулентных, а также от других микобактерий. Повторяя туберкулезную палочку по ряду свойств, имеющих отношение к болезнетворности, они лишены их полного набора. Арсенал реально действующих начал не ясен и скорее всего раскрывается лишь в конфликте с хозяином. Не исключено, что некоторые факторы образуются только внутри зараженных клеток, как это происходит у других бактерий, предпочитающих внутриклеточный образ жизни. Например, сальмонеллы, паразитируя внутри макрофагов, дополнительно экспрессируют более 40 генов.
Гранулема. Взаимодействие между туберкулезной палочкой и макрофагами инициирует базисный для туберкулеза процесс — воспаление гранулематозного типа. Сама по себе гранулема не уникальна для туберкулеза. Сходные реакции возникают при других хронических инфекциях (бруцеллез, сифилис, кокцидиоидомикоз, лейшманиоз и пр.), а также при внедрении любых трудно метаболизируемых материалов. Например, гранулемы служат основой пневмоний, развивающихся при длительном вдыхании минеральной пыли. Множественные очаги гранулематозного воспаления образуются после внутривенных инъекций зимозана (полисахарид клеточной стенки дрожжей), угольных частиц и пр. Еще Кох обратил внимание на появление гранулем после введения животным убитых туберкулезных палочек или их дериватов. Такие реакции не перерастают в процесс, так как инъецируемый материал лишен способности к самовоспроизведению.
Зарождаясь тотчас после инфицирования, туберкулезные гранулемы получают мощный импульс с появлением Т-лимфоцитов, сенсибилизированных к возбудителю. Неспецифическая (доиммунная) гранулема трансформируется в специфическую (постиммунную), обретая признаки, характерные для туберкулеза. Именно с этого момента гранулема называется туберкулом. Подчиняясь диалектике воспаления, она служит не только рычагом саногенеза, но и механизмом повреждения. Следовательно, чтобы понять туберкулезный процесс, необходимо знать факторы, побуждающие гранулему к патогенетически значимой эволюции.
Неспецифическая (доиммунная) гранулема. Первичное заражение макрофагов не проходит бесследно. Добившись права на внутриклеточную репликацию, туберкулезная палочка вызывает их повреждение — пусть незначительное, но достаточное, чтобы стать центром для притяжения моноцитов крови, которые, попав в зону инфекции, трансформируются в макрофаги. Главным инструментом первичного повреждения служат липиды клеточной стенки с мембранотоксическим эффектом. Аппарат пиогенного (нейтрофилзависимого) воспаления после краткой вспышки практически бездействует. На протяжении всего процесса нейтрофилы остаются беспомощными свидетелями развертывающихся событий, располагаясь в небольшом количестве по периферии гранулемы. Если они и проявляют активность, то это скорее приносит вред, способствуя протеолизу некротических масс, который выгоден возбудителю.
Объяснение макрофагальной доминанты следует искать в природе микобактерий. Проникая в клетки, они исчезают из поля зрения эффекторов острого воспаления (комплемента, нейтрофилов, а затем и антител), которое гаснет, не получая стимулов к развитию. Медленное размножение туберкулезных бактерий и отсутствие у них ярких деструктивных начал снижает вероятность острого конфликта и в дальнейшем.
Стимулом к воспалению служит не только ортодоксальное повреждение клеток. Компоненты клеточной стенки (микозиды, сульфатиды, корд-фактор, липоарабиноманнан) побуждают макрофаги к секреции цитокинов и других флогогенных молекул, которые, привлекая моноциты, усиливают эффекторные ресурсы гранулемы и дирижируют событиями внутри нее. Кажется странным, но вирулентные штаммы в этом отношении менее активны. Например, их липоарабиноманнан не только не возбуждает, но даже блокирует секреторную реакцию макрофагов. Однако это формальное противоречие. Задержка воспалительного процесса способствует опережающему размножению бактерий, невольно содействуя стабилизации инфекта. Имеет значение и то, что туберкулезная палочка (тот же липоарабиноманнан), снижая чувствительность макрофагов к гамма-интерферону, ослабляет HLA-зависимое представление антигенов и пролиферацию Т-лимфоцитов.
Так или иначе, но доиммунная гранулема не останавливает инфекцию. Из легких туберкулезная палочка проникает в регионарные лимфатические узлы и далее в кровоток, получая возможность для диссеминации. Этому содействует сродство к макрофагам, присутствующим во всех тканях. За редким исключением дело ограничивается преходящими гранулематозными (в терминологии фтизиатров — параспецифическими) реакциями, повторяющими картину первичного очага инфекции. Сама по себе бактериемия на этом этапе не опасна, но она создает условия для персистенции возбудителя и его отсроченной реактивации в различных органах. С клинической точки зрения доиммунная фаза соответствует инкубационному периоду, так как никаких реальных симптомов здесь не возникает.
Специфическая (постиммунная) гранулема. Стратегическим итогом неспецифической гранулемы является развитие иммунного ответа против туберкулезной палочки. Реакция стартует в регионарных лимфоузлах, куда возбудитель и его антигены транспортируются (макрофагами, дендритными клетками) из первичной гранулемы. Дальнейшие события тесно связаны со специфическим воспалением, в основе которого лежит аллергическая реакция на микобактериальные антигены. Представления об аллергии как о необычной чувствительности к обычным (безвредным для нормального, т.е. несенсибилизированного организма) антигенам зародились именно при изучении микобактерий туберкулеза.
Все началось с наблюдений Коха. Он обнаружил, что у инфицированных животных чувствительность к туберкулезной палочке и ее продуктам гораздо выше, чем в контроле. Феномен Коха сводится к следующему. При подкожном заражении здоровой морской свинки культурой туберкулезной палочки видимые изменения на месте инъекции в первые дни отсутствуют. Лишь спустя 10—14 дней возникает плотный, изъязвляющийся узелок, который не заживает до гибели животного от генерализованного туберкулеза. Но если туберкулезные бактерии (живые, убитые) или их дериваты ввести внутрикожно или подкожно зараженному животному, то воспалительная реакция возникает уже через 24—48 ч. Ей сопутствуют общая интоксикация и обострение отдаленных очагов.
Отсроченный ответ при заражении здорового организма объясняется тем, что бактерии, которые размножаются в зоне инокуляции, до определенного момента сохраняют патогенетическую инертность. Ее нарушает сам хозяин, который после иммунной перестройки провоцирует гиперергическую (аллергическую) реакцию против возбудителя. В этом повинны сенсибилизированные Т-лимфоциты, круто меняющие ход событий. Они превращают доиммунную гранулему в мощный эффекторный аппарат, наделяя реакцию морфологией и агрессивностью истинного туберкула. Гранулема увеличивается в размерах за счет обильного притока новых клеток и объединения соседних бугорков. В ней появляются гигантские многоядерные клетки Лангерганса—Пирогова (результат слияния активированных макрофагов), макрофаги трансформируются в эпителиоидные клетки, центр гранулемы подвергается казеозному (творожистому) некрозу, по периферии накапливаются фибробласты, побуждаемые к секреции коллагена и отграничению зоны конфликта соединительнотканной (фиброзной) капсулой (рис. 4). Зрелый туберкул выглядит как плотный, сероватого (позднее желтоватого) цвета узелок диаметром 1—3 мм, не вылущиваемый из окружающей ткани.

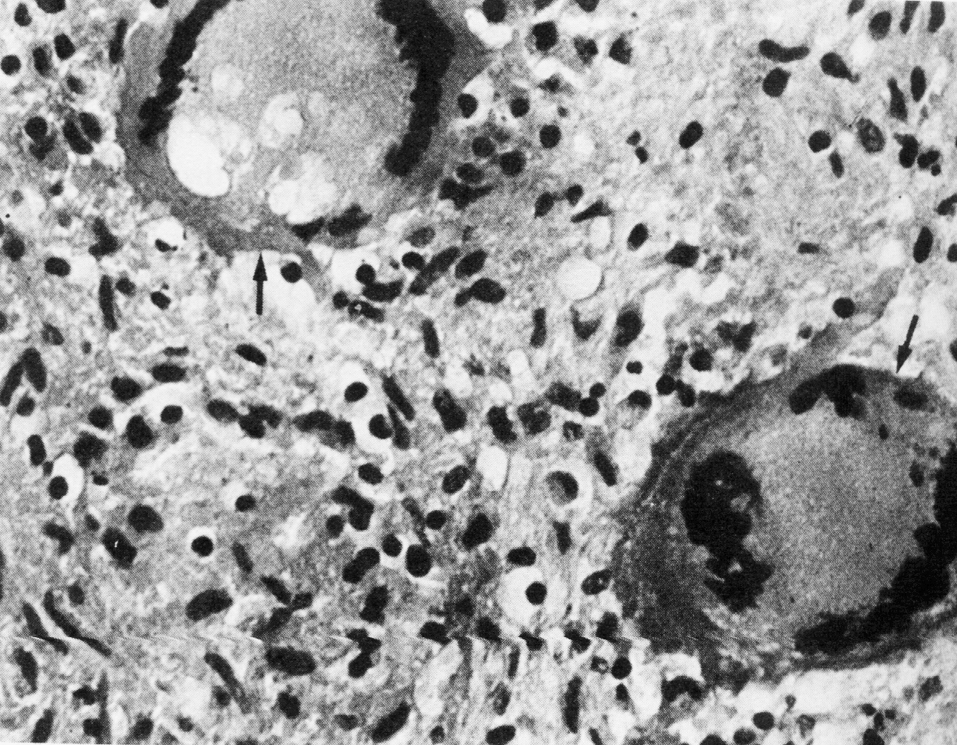
Суть этой феноменологии состоит в Т-зависимой активации макрофагов, которые повышают свою агрессивность против поглощенных бактерий и начинают интенсивно секретировать флогогенные цитокины и другие медиаторы с широким спектром местных и дистантных эффектов. Приток Т-лимфоцитов, сенсибилизированных против туберкулезных антигенов, знаменует кризис воспалительной реакции и фактически провоцирует ее обострение, выводя из двусмысленного (т.е. весьма ненадежного) гомеостаза. Среди Т-цитокинов на главную (но не единственную) роль претендует гамма-интерферон, который потенцирует гибель внутримакрофагальных бактерий. Участие гамма-интерферона в защите человека от микобактерий подтверждают наблюдения о повышенной склонности к туберкулезу и микобактериозам детей с генетическими дефектами гамма-интерфероновых рецепторов.
Для уничтожения поглощенных бактерий стимулированные макрофаги применяют тактику активной и пассивной (если такое словосочетание возможно!) агрессии. В первом случае действуют факторы и механизмы с прямым бактерицидным и бактериостатическим эффектом — реактивные метаболиты кислорода и азота, сокращение лабильного пула железа и пр. Пассивная агрессия связана с гибелью самих макрофагов, которую вызывают цитотоксические Т-лимфоциты, атакующие зараженные клетки. Это лишает туберкулезную палочку удобной среды обитания.
Но клетки гранулематозного инфильтрата гибнут и по другой, менее элегантной, но более значимой причине. Речь идет о казеозном распаде, связанном с недостатком кровоснабжения в быстро растущей гранулеме. Этот парадокс символизирует начало обратного развития событий, так как в казеозной зоне большинство микобактерий гибнет, не выдерживая аноксии, закисления среды и недостатка факторов питания (прежде всего железа). Массовая гибель бактерий радикально снижает концентрацию болезнетворного начала, обрывая воспалительную реакцию. Активированные фибробласты секретируют коллаген и, репарируя очаг повреждения, завершают саногенез. Нередко это сочетается с кальцификацией (обызвествлением) туберкула.
Описанные события — результат сбалансированного взаимодействия клеток, тонко чувствующих нарушения тканевого гомеостаза. Специфическая гранулема действует как рычаг иммунитета, от надежности которого зависит вероятность самоизлечения. Ослабление гранулематозной реакции снижает резистентность к возбудителю, содействуя эволюции туберкулезного процесса. Дождавшись сигнала (точнее сигналов) от Т-лимфоцитов, макрофаги сами становятся мощным источником цитокинов и других медиаторных молекул, определяющих события внутри гранулемы. Одна из центральных позиций принадлежит альфа-туморонекротическому фактору (ТНФ-a), главным источником которого служат активированные макрофаги. В опытах на животных нейтрализация ТНФ-a (антителами или растворимыми рецепторами) подавляет рост гранулем, повышая чувствительность к туберкулезной инфекции и провоцируя ее осложнения. Это означает, что ТНФ-a поддерживает непрерывное обновление клеточного состава гранулем, сдвигая баланс в пользу хозяина. Кстати, гипоксия, вносящая решающий вклад в казеозный распад туберкулов, обусловлена не только отставанием ангиогенеза от быстро растущей клеточной массы, но и цитокининдуцированным (прежде всего тем же ТНФ-a) повышением тромбогенности эндотелия микрососудов.
В любом случае эффекторные реалии гранулемы ощутимы лишь с приходом сенсибилизированных Т-лимфоцитов. Принципиально, что рост постиммунной гранулемы сочетается с усилением антибактериальной активности макрофагов. Этого явно не хватает неспецифической воспалительной реакции. Поэтому Т-лимфоциты следует рассматривать не только как фактор количественного усиления процесса, но и как стимул для его перехода в новое качество — механизм, одинаково важный для сано- и патогенеза.
Деструкция. На фоне безуспешных поисков специфических токсинов уже в начале XX столетия было постулировано, что болезнетворность туберкулезной палочки связана не с прямым повреждением тканей, а с длительным выживанием бактерий в организме. Именно к этому свелось большинство исследований по изучению патогенеза туберкулеза.
Как уже говорилось, подобно любой воспалительной реакции, туберкулезная гранулема, исходно нацеленная на саногенез, рождает повреждение с зачатками клинически значимой патологии. Туберкул, эволюционирующий в направлении саногенеза, прорастает фиброзной тканью, т.е. превращается в рубец. Такой исход знаменует успешное завершение борьбы с возбудителем, стартовавшей на доиммунном этапе гранулематозного процесса. Но иногда некроз прогрессирует, а казеозная масса вместо рубцевания подвергается расплавлению протеолитическими ферментами макрофагов и, возможно, нейтрофилов. В такой среде туберкулезная палочка чувствует себя великолепно и, размножаясь, поддерживает альтерацию. Если туберкул, переживающий фазу распада, находится по соседству с бронхами или кровеносными сосудами, они втягиваются в некротический процесс. В подобных случаях возбудитель получает возможность для распространения на другие участки легкого, а попав в циркуляцию, вызывает обсеменение отдаленных органов с тяжелейшими последствиями. О причинах, сдвигающих баланс в сторону патогенеза, можно только гадать, но они, безусловно, связаны не с прямым действием бактерий, а опосредованы иммунологическими механизмами.
Интоксикация. Интоксикация — одно из обязательных проявлений туберкулеза. Она носит неспецифический характер, включая такие симптомы как лихорадка, потливость, ночные поты, ознобы, быстрая утомляемость, слабость, тахикардия, снижение аппетита, исхудание и пр. Истощение может обретать крайние формы, оправдывая историческое название болезни — чахотка (греч. phthisis — истощение, англ. consumption — потребление, истощение). Симптомы нарастают вместе с туберкулезным процессом, но иногда возникают без очевидной органной патологии. В клинических классификациях туберкулезная интоксикация («туберкулез без определенной классификации») выделена в особую форму туберкулеза детей и подростков.
Интоксикацию удается воспроизвести на животных при помощи липидных и гликолипидных фракций клеточной стенки микобактерий, в частности, того же корд-фактора. Но не это главное. Поиски туберкулезных токсинов привели к неожиданному, но вполне логичному открытию, которое следовало из наблюдений о повышенной чувствительности инфицированного организма к туберкулезной палочке. Путем выпаривания водно-глицериновой вытяжки из культур М. tuberculosis Кох приготовил препарат (так называемый старый туберкулин Коха), безвредный для нормальных, но токсичный для животных, зараженных микобактериями туберкулеза. Его действующим началом оказались белковые дериваты туберкулезной палочки. Внутрикожное или подкожное введение туберкулина вызывало не только местную воспалительную реакцию, но и общую интоксикацию с обострением очагов туберкулезной инфекции. То же самое наблюдалось у туберкулезных больных, вынудив приостановить увлечение туберкулинотерапией. Но препарат оказался полезным в диагностике и с этой целью его стали применять уже при жизни Коха.
Сегодня мало кто сомневается, что туберкулезная интоксикация носит опосредованный характер, отражая гиперпродукцию цитокинов клетками гранулемы, прежде всего активированными макрофагами. Это, в частности, справедливо для одного из кардинальных признаков хронического туберкулеза — кахексии. Ее связывают с действием ТНФ-a, который усиливает липазную активность адипоцитов, заслужив репутацию кахектина. Отсюда с патогенетической точки зрения местные повреждения и общую интоксикацию следует считать однотипными процессами. Они базируются на событиях, которые происходят внутри гранулемы и сопровождаются повышением ее местной и общей агрессивности.
Персистенция и реактивация. С микробиологической точки зрения выздоровление от туберкулезной инфекции не бывает полным. Пусть в ничтожном количестве, но возбудитель сохраняет жизнеспособность и потенциальную агрессивность в законсервированных гранулемах. Это было осознано еще современниками Коха после опытов с заражением морских свинок материалом из «бывших» туберкулов.
Латенция связана со способностью туберкулезной палочки длительно выживать в метаболически инертном состоянии, реализуя широкие способности к адаптивной изменчивости. Допускается, например, возможность трансформации в L-формы, что в известном смысле стыкуется с представлениями о фильтрующихся и других атипичных вариантах микобактерий туберкулеза. Обсуждается феномен так называемых некультивируемых бактерий, которые неопределенно долго сохраняют жизнеспособность без репликации, т.е. без того, что считалось обязательным для бесспоровых бактерий. Согласно еще одной версии, причиной латенции может быть изоляция туберкулезной палочки от ресурсов питания в связи с функциональными изменениями ее плазматической мембраны. Так или иначе, но следует признать, что рост и размножение — необязательное условие для выживания возбудителя туберкулеза в инфицированном организме. После завершения бурной реакции хозяин обычно сохраняет внедрившиеся бактерии на всю жизнь. Это поддерживает опасность эндогенных рецидивов, склоняя к парадоксу: гранулема не умирает с завершением процесса, продолжая жить после собственной смерти.
Не менее загадочен и механизм реактивации туберкулезной инфекции. К этому предрасполагают истощение организма (особенно белковое голодание), хронические воспалительные процессы органов дыхания, диабет, акоголизм и пр. Из-за отсутствия конкретных улик приходится говорить о снижении резистентности, ослаблении иммунитета, что, по сути, не более, чем декларации. Повышенная склонность к туберкулезу при нарушениях иммунитета (в частности, у больных, инфицированных вирусом иммунодефицита человека — ВИЧ) в известной мере проясняет картину, но не является аналогом того, что происходит в других ситуациях.
Следует учитывать состояние не только общего иммунитета, но и резистентность органа-мишени (легкие), где возникает реактивация. Очевидно, что зажившие гранулемы (даже кальцификаты) нельзя считать мертвой зоной. Возбудитель сохраняет здесь потенциальную активность, требуя от хозяина хотя бы минимального напряжения гомеостатических ресурсов. Не исключено, что латентная инфекция служит перманентным источником антигенов, тонизирующих иммунитет, который при туберкулезе принято считать нестерильным. Это косвенно подтверждается и динамикой постинфекционной аллергии к туберкулину: она имеет тенденцию не только к длительному сохранению, но часто усиливается перед рецидивом заболевания. Складывается впечатление, что механизм, предназначенный для сдерживания инфекта, провоцирует реанимацию возбудителя.
Впрочем, активация возможна и без каких-либо очевидных причин, на фоне физического и социального благополучия. Это побуждает к размышлениям о генетических факторах и наследственной предрасположенности к туберкулезу. Конкретные выводы по этому поводу отсутствуют, хотя в опытах на линейных животных чувствительность к туберкулезной инфекции отчетливо сопряжена с генетическим фоном. Есть указания на то, что устойчивость к туберкулезу ассоциирована с HLA-фенотипом, но и здесь пока мало определенности. Вызывают интерес (но не могут быть основой для обобщений) наблюдения о гипорезистентности к возбудителям туберкулеза и микобактериозов у лиц с избирательными иммунодефицитами, например с врожденным дефектом по Th1-цитокинам и их рецепторам (гамма-интерферон, интерлейкин-12).
Иммунитет и аллергия. Человек высоковосприимчив к туберкулезной палочке. Широкое (в прошлом почти поголовное) распространение аллергии к туберкулину говорит о том, что заражение обычно запускает процесс, который сопровождается иммунным ответом. Вместе с тем, из массы зараженных людей туберкулезом болеют немногие.
Главным или даже единственным эффектором приобретенного противотуберкулезного иммунитета являются Т-лимфоциты. Антитела скорее всего лишены протективности, хотя в прошлом не раз сообщалось о лечебном действии сывороток животных, иммунизированных микобактериями туберкулеза и их дериватами. Это оказалось иллюзией, отвергнутой временем, но попытки реанимировать идею о гуморальном противотуберкулезном иммунитете (в частности, на основе представлений о функциональной неоднородности антител) продолжаются и сегодня.
Туберкулезная палочка знаменита свои мощным иммуноадъювантным эффектом, которым давно пользуются для усиления иммунных реакций. Это важнейший признак микобактерий, от которого во многом зависит развитие туберкулезного процесса. Вакциной БЦЖ (см. ниже) пытались даже лечить больных злокачественными опухолями, но дальше надежд и не очень убедительных результатов дело не пошло. Поначалу адъювантность связывали с липидными фракциями, однако в чистом виде они не эффективны. Позже акцент был смещен в пользу пептидогликана (точнее, его мурамилдипептидного фрагмента), но в этом случае трудно понять уникальность иммуноадъювантности микобактерий.
Туберкулезная инфекция принадлежит к классике иммунологии, будучи эталоном одной из разновидностей аллергии — гиперчувствительности замедленного типа (ГЗТ). Базисное открытие принадлежит Коху, обнаружившему повышенную реактивность к туберкулину у инфицированных животных. Лечение туберкулином больных туберкулезом, которое с подачи Коха увлекло было его современников, быстро сменилось разочарованием, так как нередко обостряло процесс, вызывая тяжелые осложнения. Для описания этого явления К. Пирке в 1906 г. ввел понятие «туберкулиновая аллергия», которая до недавних пор оставалась синонимом ГЗТ. Современный этап ее изучения стартовал в конце 1940-х гг., когда М. Чейз и его коллеги установили возможность переноса аллергии к туберкулину при помощи лимфоцитов, полученных от сенсибилизированных животных. Эта работа стала основой современных представлений о клеточном иммунитете и иммунопатогенезе.
Реакция на туберкулин — не что иное, как очаг гранулематозного воспаления, спровоцированного взаимодействием между антигенами туберкулезной палочки и сенсибилизированными Т-лимфоцитами. Постиммунную гранулему можно рассматривать как вынужденную меру — гиперболизированное проявление ГЗТ в ответ на внедрение антигенного объекта, не поддающегося уничтожению в масштабе обычного ответа. Как уже говорилось, иммунологически зависимое усиление гранулематозной реакции необходимо для подавления туберкулезной инфекции. Но ясно и то, что, если деструкция опережает репарацию, акцент смещается в сторону патогенеза, часто обретая необратимый характер. Это характерно для вторичного туберкулеза, в основе которого лежит хроническое повреждение тканей на фоне реактивации персистентной туберкулезной инфекции. Формируется порочный круг, когда, укрепляя гематотканевой барьер и препятствуя распространению инфекции, зона гиперергического некроза превращается (после расплавления казеозных масс) в отличную среду для размножения туберкулезных бактерий. Т-зависимая аллергия становится инструментом клинически значимого повреждения, а безобидные туберкулопротеины (действующее начало туберкулина) превращаются в лидирующий фактор болезнетворности. Казалось бы, можно остановить процесс, подавив агрессивность гранулемы, но это ослабит сопротивление возбудителю и обострит заболевание. Нечто подобное имеет место при ВИЧ-инфекции на фоне ослабления Т-клеточного иммунитета. Реактивационный (вторичный) туберкулез у таких больных протекает с признаками генерализации, характерными для первичной инфекции, иногда без типичных гранулематозных реакций.
Взаимоотношения между протективным иммунитетом и аллергией при туберкулезе до сих пор вызывают споры. Интрига вокруг их возможной разобщенности недавно получила очередной допинг в связи с развитием представлений о гетерогенности Т-лимфоцитов. Оказалось, что традиционное подразделение на Т-хелперы (CD4+) и Т-киллеры (CD8+) не исчерпывает эффекторного репертуара Т-лимфоцитов: и те и другие функционально не однородны. По профилю секретируемых цитокинов дифференцируют, как минимум, два варианта Т-хелперов (Тh1 и Th2), которые различаются по активности в реакциях клеточного и гуморального иммунитета. Тh1 управляют стимуляцией макрофагов (через гамма-интерферон и др.) и дифференцировкой цитотоксических CD8+ Т-клеток (интерлейкин-2 и др.); Th2 дирижируют реакциями В-лимфоцитов (интерлейкины-4, 5 и др.). Кроме того, продуцируя ингибиторные цитокины, Тh1 и Th2 взаимно контролируют друг друга, оправдывая представления о Т-зависимой иммуносупрессии (дополнительные супрессорные сигналы исходят из субпопуляций CD8+ клеток).
В аллергии к туберкулезной палочке задействованы главным образом Тh1-хелперы. Секретируя макрофаготропные цитокины, они повышают антимикробную (к сожалению, и антитканевую) агрессивность макрофагов и гранулематозных реакций в целом. Генетические дефекты по Тh1-цитокинам снижают устойчивость к туберкулезной палочке и возбудителям оппортунистических микобактериозов. Этому сопутствует ослабление гранулематозных реакций, которые мобилизуют слишком мало макрофагов, чтобы противостоять инфекции.
Цитотоксические CD8+ лимфоциты больше нацелены на прямое уничтожение зараженных клеток, экспрессирующих микробные антигены в комплексе с HLA-1. Действенность этого механизма должна подкрепляться гибелью внутриклеточных микробов. Такой финал логичен при вирусных инфекциях (смерть клетки-хозяина обрывает репликативный цикл вируса), но менее очевиден для оказавшихся на свободе бактерий: часть из них сохраняет жизнеспособность и готовность к заражению соседних клеток. Безусловно, активированные макрофаги агрессивнее клеток доиммунной гранулемы, но, во-первых, не все из них одинаково чувствительны к Т-цитокинам, разновременно включаясь в деятельное сопротивление бактериям, а, во-вторых, туберкулезная палочка способна выживать в активированных макрофагах. Не спасает и то, что Т-лимфоциты секретируют факторы, непосредственно убивающие микобактерии. В гранулах CD8+ Т-лимфоцитов содержится сапониноподобный белок гранулизин, который убивает М. tuberculosis, действуя на липиды клеточной стенки. Гранулизин активен не только вне клеток, но и впрыскивается внутрь зараженных макрофагов. Вклад такого рода веществ в реальную протективность не известен.
Идеи о функциональной неоднородности Т-лимфоцитов обрели подкрепление после открытия неклассических антигенпредставляющих молекул, обособленных от молекул главного комплекса гистосовместимости. Они относятся к семейству CD1-рецепторов и в отличие от HLA, презентирующих пептиды, экспрессируют липидные и гликолипидные антигены. Т-лимфоциты, распознающие эпитопы в комплексе с CD1 (они получили название Т-естественных киллеров), выявлены среди CD8+ Т-лимфоцитов, а также в минорной фракции Т-клеток, негативных по базисным Т-маркерам. CD1-зависимые Т-лимфоциты, настроенные против микобактериальных липидов и гликолипидов, продуцируют макрофаготропные и другие провоспалительные цитокины и в этом отношении похожи на Тh1. Некоторые из них не только лизируют зараженные клетки, но и секретируют факторы, самостоятельно убивающие бактерии.
CD1-зависимая кооперация расширяет масштаб взаимодействия иммунокомпетентных клеток с микробными агентами. Не исключено, например, что стимуляция гликолипидными антигенами извращает цитокиновый баланс в лимфоидной ткани, создавая условия для гиперактивации определенных субпопуляций Т-лимфоцитов. Возможно, именно за этим скрывается причина высокой иммуноадъювантности туберкулезной палочки и гиперболизации Th1-опосредованных реакций. Благодаря CD1 в презентацию антигенов втягиваются клетки, слабо экспрессирующие HLA-1 и HLA-2. Это может иметь определенные последствия тем более, что туберкулезные микозиды снижают количество HLA на поверхности антигенпредставляющих клеток.
Обобщая, туберкулез следует отнести к реактивным (иммунологически зависимым) патологиям, возбудители которых, не обладая достаточным арсеналом прямых деструктивных начал, поддерживают болезнь за счет конфликта с эффекторами иммунитета, вынуждая их к патогенетически значимым реакциям (рис. 5).


Патология
Клинический спектр туберкулезной инфекции необычайно широк — от малозаметных проявлений до опасных для жизни заболеваний, от неспецифической интоксикации до вполне конкретных органных нарушений, от острых и скоротечных симптомов до хронической патологии. Чаще всего страдают легкие, но от поражений не застрахован ни один орган.
При всем клиническом разнообразии принято выделять два патогенетических варианта туберкулеза — первичный и вторичный. Первичный туберкулез возникает у лиц, ранее не имевших контакта с возбудителем. Это означает, что взаимоотношения с туберкулезной палочкой здесь не зависят от исходного иммунологического фона (туберкулезного анамнеза), а определяются особенностями врожденного иммунитета. Реакции специфического иммунитета включаются с опозданием, но успевают существенно повлиять на течение процесса и клиническую симптоматику. Что касается вторичного туберкулеза, то его без натяжки можно отнести к категории иммунопатологий.
Принципиальная схема взаимоотношений человека с М. tuberculosis приведена на рис. 6.

Первичный туберкулез. Вероятность контакта с туберкулезной палочкой зависит от эпидемической ситуации. В годы широкого распространения туберкулеза (первые 20—30 лет XX столетия) инфицирование обычно происходило в раннем детстве или подростковом возрасте. К моменту половой зрелости более 90% населения было заражено М. tuberculosis. Такая тенденция и сегодня сохраняется в регионах, эндемичных по туберкулезу. Напротив, резкое сокращение заболеваемости в развитых странах привело к значительному снижению этого показателя и относительному повышению числа первично инфицируемых взрослых людей.
Клинические проявления первичного туберкулеза возникают у 3—5% инфицированных в течение года после заражения. В зоне внедрения (обычно легкие) возбудитель захватывается макрофагами, возбуждая воспалительную реакцию (неспецифическая гранулема). Бактерии легко проходят этот барьер, быстро проникают в регионарные лимфатические узлы и далее, через кровяное русло, в отдаленные органы. Развитие истинного туберкула (т.е. специфической гранулемы) занимает 2—3 нед и связано с подключением сенсибилизированных Т-лимфоцитов. Формируется первичный аффект, а втягивание в процесс регионарной лимфоидной ткани ведет к лимфадениту. Увеличение трахеобронхиальных и медиастинальных лимфоузлов типично для первичного туберкулеза и может быть его единственным проявлением.
Первичный аффект в сочетании с регионарным лимфаденитом называется первичным туберкулезным комплексом. Чаще всего он самоизлечивается, подвергаясь фиброзу и кальцификации, и выявляется ретроспективно при случайных рентгенологических осмотрах (кальцифицированные очаги Гона в легких). Но возможны и серьезные осложнения. Казеозное разъедание бронха грозит распространением инфекции на легочную ткань с развитием опасной для жизни диссеминированной бронхопневмонии и эмпиемы плевры. Туберкулезные очаги в лимфатических узлах, переживающие казеозный распад, иногда прорываются в перикард или, увеличиваясь в размере, сдавливают крупные бронхи, вызывая ателектаз сегмента и даже целой доли легкого. Гранулематозный процесс может повредить стенку прилегающих сосудов, открывая путь в циркуляцию громадному числу бактерий. Они запускают тяжелейшую форму генерализованной инфекции — милиарный туберкулез (лат. milium — просяное семя), когда во многих органах образуется изобилие мелких (1—2 мм) туберкулов. Милиарный туберкулез чаще наблюдается у детей первых трех лет жизни и без лечения убивает в течение месяца. Смерть наступает от менингита, причем в очагах поражения количество микобактерий ничтожно (иммунопатогенез?!).
Впрочем, проявления системной инфекции не всегда столь катастрофичны. Более того, они обычно отсутствуют. Бактериемия имеет скорее стратегическое значение, закладывая очаги персистентной инфекции, потенциально готовые к реактивации. Однако набрав ход, процесс течет быстро и необратимо, за что получил название острого туберкулеза в противовес вторичному (хроническому) туберкулезу, который без адекватной терапии медленно, но неизбежно ведет к смерти.
Вторичный туберкулез. Вторичный туберкулез обычно возникает при реактивации эндогенного фокуса примерно у 10% инфицированных через пять и более лет после первичного заражения. Реинфицирование извне тоже возможно, но наблюдается реже. По данным сравнительного генотипирования выделенных штаммов, заражение извне наблюдается в 20—44% всех случаев вторичного туберкулеза.
Вторичный туберкулез поражает детей и взрослых, хотя в прошлом был известен как туберкулез взрослого типа. Сегодня, когда первичное заражение взрослых случается относительно часто, понятия «взрослого» и «детского» туберкулеза потеряли смысл, хотя и акцентируют внимание на особенностях туберкулезного процесса в сенсибилизированном и нормальном организме.
Склонность к вторичному туберкулезу повышается при неблагоприятных условиях жизни, на фоне хронических болезней, стрессов, но в целом причины оживления инфекции расплывчаты, и вероятность вторичного туберкулеза практически непредсказуема. Пожалуй, единственным исключением является фон, создаваемый ВИЧ-инфекцией: среди туберкулинопозитивных больных синдромом приобретенного иммунодефицита туберкулез встречается в 500 раз чаще, чем в общей популяции.
Вторичный туберкулез протекает как хроническое заболевание, которое не излечивается без этиотропной (антимикробной) терапии. Почти всегда это пульмонарная патология, которая начинается коварно, чаще с верхушки правого легкого. В зоне реактивации возникает конгломерат растущих туберкулов, которые, сливаясь, образуют крупный очаг казеозного распада. Подвергаясь протеолитическому разжижению, казеозная масса прорывается через бронх с образованием полости — каверны. Со временем каверна окружается плотной фиброзной тканью, препятствующей ее сжатию и заживлению (рубцеванию). Разъедание сосудов ведет к кровохарканью и легочным кровотечениям.
Благодаря обильной аэрации и отличному питательному материалу (продукты протеолиза) туберкулезная палочка получает условия для бурной внеклеточной репликации. Накапливаясь в огромных количествах по внутреннему периметру кавернозной стенки, бактерии поддерживают агрессивность местного процесса, а, выделяясь с мокротой, превращают больного в источник инфекции. Такие случаи (они более типичны для взрослых, чем для детей и подростков) называют открытым туберкулезом.
Распространение на регионарные лимфатические узлы и лимфогематогенная диссеминация не типичны. Это связано с закупоркой лимфатических сосудов и капилляров некротическими массами в результате интенсивной воспалительной реакции, которая возникает на фоне аллергии к туберкулезной палочке и по сути повторяет модельный феномен Коха. Впрочем, генерализация все же происходит, хотя и иначе, чем при первичном туберкулезе. Выход высококонтагиозного материала в воздухоносные пути создает предпосылки для распространения бактерий на легочную ткань с развитием новых каверн и даже фатальной бронхопневмонии (скоротечная чахотка). Кроме того, с мокротой возбудитель попадает в гортань, ротовую полость и кишечник, угрожая внепульмонарными осложнениями.
Бактериологический диагноз
Бактериоскопия и другие некультуральные методы. Лучшим материалом для выявления туберкулезной палочки служит мокрота. Кроме того, в зависимости от локализации процесса исследуют мочу, кал, спинномозговую жидкость, экссудат из полостей, биоптаты тканей.
Открытый туберкулез легких — одна из немногих инфекций, при которых чрезвычайно полезна микроскопия материала, полученного от больного. Это классический образец экспресс-диагностики — микробиологического анализа без выделения чистой культуры. Здесь он строится на тинкториальном своеобразии возбудителя, а именно на его устойчивости к обесцвечиванию кислотами. Кроме окраски по Цилю—Нильсену, используют флюорохромные красители (аурамин, родамин), исследуя препараты под малым увеличением люминесцентного микроскопа. Это позволяет быстро просмотреть большое число полей зрения и тем самым повысить чувствительность метода.
И все-таки без дополнительных ухищрений разрешающая способность бактериоскопии довольно низка: в 1 мл должно быть не меньше 100000 микробных тел. Чтобы сделать метод более надежным, бактерии концентрируют, т.е. повышают их содержание в единице объема. Для этого мокроту, обработанную щелочью, подвергают центрифугированию или флотации (после добавления ксилола/бензола и тщательного встряхивания микобактерии, богатые липидами, всплывают вместе с пеной). Можно поступить проще, сделав толстый мазок путем последовательного наслоения-подсушивания нескольких капель мокроты. Используя эти приемы, положительный результат удается получить при содержании 5000—10000 бактерий в 1 мл мокроты. Это значительно ниже чувствительности культурального метода, но компенсируется быстротой, доступностью, а главное — эпидемиологической значимостью анализа. Дело в том, что положительный результат говорит о массивном выделении возбудителя, отражая опасность больного для окружающих. По этому признаку туберкулезных больных делят на бациллярных и абациллярных, подразумевая, что первые требуют особенно пристального внимания. Этиотропная терапия, снимающая бациллярность, делает больного практически незаразным.
Неоднократно делались попытки заменить или хотя бы усилить бактериоскопию другими методами экспресс-диагностики. С этой целью испытывались различные способы выявления антигенов и специфических метаболитов туберкулезной палочки. За немногими исключениями (например, определение туберкулостеариновой кислоты в спинномозговой жидкости и мокроте) они не принесли удовлетворения из-за плохой воспроизводимости или малой чувствительности. Одно из новейших направлений в диагностике туберкулеза основано на выявлении генетических маркеров (специфических последовательностей ДНК) М. tuberculosis в полимеразной цепной реакции (ПЦР). Чувствительность современных вариантов ПЦР составляет 70—90% от культурального метода, дает положительный результат в 90—95% образцов мокроты от бациллярных больных и позволяет подтвердить диагноз у 50—60% абациллярных больных. Одна из технических трудностей ПЦР связана с ингибиторами ДНК-полимеразы, которые могут присутствовать в исследуемом материале.
Культуральные методы. Максимальной чувствительностью (если не считать заражения мокротой морских свинок — способ, который сегодня с диагностическими целями не используется) обладают культуральные методы, основанные на выделении и идентификации чистой культуры туберкулезной палочки. Они позволяют обнаружить 20—100 бактерий в 1 мл исследуемого материала. Для подавления сопутствующей микрофлоры мокроту деконтаминируют, пользуясь устойчивостью микобактерий к кислотам, щелочам и дезинфектантам. Обработанную пробу засевают на специальные питательные среды и инкубируют не менее 8 нед, наблюдая за динамикой роста. Культуры идентифицируют по характерным свойствам, спектр которых достаточно широк — от культуральных и морфотинкториальных признаков до профиля миколовых кислот и генетических маркеров. Неспособность к пигментообразованию и медленный рост позволяют отличить М. tuberculosis от большинства нетуберкулезных микобактерий. Позитивный ниациновый тест (в нем учитывается накопление в культуре ниацина, т.е. никотиновой кислоты) дифференцирует М. tuberculosis от М. bovis и беспигментных медленнорастущих микобактерий. Редкие ниациннегативные изоляты М. tuberculosis можно распознать по восстановлению нитратов.
Большинство штаммов М. tuberculosis вырастают в течение 4 нед, но возможна отсрочка (до 12 нед и более), особенно при исследовании материала от больных, леченных антитуберкулезными препаратами. Благодаря высокой чувствительности культуральные методы остаются золотым стандартом, т.е. точкой отсчета для суждения о разрешающей способности других методов. Но возможны и парадоксы, когда отрицательный анализ сочетается с позитивными результатами бактериоскопии мокроты, а тем более ПЦР. Это связано с гибелью микобактерий при деконтаминации или с утратой способности к размножению под влиянием антибиотиков. Первую причину удается сгладить, подняв специфичность деконтаминирующих процедур. Для этого пользуются мягким обеззараживанием мокроты или культивируют ее в присутствии антибиотиков, которые подавляют размножение других бактерий, не влияя на рост туберкулезной палочки.
Принципиальный недостаток культуральных методов — медленный рост микобактерий туберкулеза и отсюда запоздалые результаты. С этим предложено бороться при помощи остроумного метода микрокультур (метод Прайса). Его суть в том, что мазки, приготовленные и подсушенные на предметных стеклах (как для микроскопии), на несколько минут погружают в 5% раствор серной кислоты (деконтаминация) и после отмывания культивируют в жидкой питательной среде. Микроколонии, выявляемые при окраске по Цилю—Нильсену, достигают максимальных размеров через 7—10 дней, хотя нередко видны уже через 3—4 сут. Есть даже предложение (впрочем, спорное) судить по виду микрокультур о болезнетворности туберкулезной палочки: вирулентные штаммы, обладающие корд-фактором, образуют змеевидные микроколонии типа жгутов/кос (см. рис. 3), авирулентные — растут в виде аморфных скоплений. Метод Прайса и его модификации не стали стандартной процедурой, так как лишены точности классического культурального анализа.
Предлагались и более изощренные способы ускоренной идентификации растущих культур, основанные, например, на измерении радиоактивного углекислого газа (14СО2), который высвобождается микобактериями туберкулеза из меченой пальмитиновой кислоты. Внедрение подобных методов в широкую практику остается проблематичным, так как сопряжено с немалыми техническими трудностями и расходами.
Иммунологическая диагностика
В 1906 г. Пирке, опираясь на наблюдения повышенной реактивности к туберкулезной палочке зараженного организма, предложил использовать кожные тесты с туберкулином для диагностики туберкулеза, точнее, туберкулезной инфекции. Для характеристики чувствительности к туберкулину Пирке ввел понятие об аллергии и ее полярном варианте — анергии. Из многочисленных модификаций «старого туберкулина Коха» закрепился препарат «очищенных белковых дериватов» — ППД (англ. PPD — purified protein derivatives). Это комплекс освобожденных от питательной среды бактериальных белков, которые в сумме представляют собой «аллерген туберкулезной палочки» — действующее начало туберкулина. Это означает, что туберкулин — не мономолекулярный субстат, а смесь микробных антигенов, которые после достаточно жесткой (денатурирующей) обработки сохраняют способность выявлять ГЗТ к микобактериям туберкулеза.
Проба Пирке предусматривала введение препарата через скарифицированную кожу. Позже (в 1910 г.) для повышения точности метода туберкулин стали вводить внутрикожно (проба Манту). Препарат дозируют в туберкулиновых единицах (ТЕ). В одной ТЕ ППД содержится 0,00006 мг сухой массы туберкулина. Обычная дозировка составляет 2 ТЕ. Ее вводят в объеме 0,1 мл в ладонную поверхность предплечья, результат учитывают по диаметру инфильтрата через 72 ч. Отсчет ведется c 5 мм, папулы меньшего размера (2—4 мм) считают сомнительными. Различают слабую (5—9 мм), среднюю (10—14 мм) и выраженную (15—16 мм) аллергию. Начиная с 17 мм у детей и подростков и с 21 мм у взрослых, реакции расценивают как гиперергические. При появлении везикулы, лимфангиита или регионарного лимфаденита реакции считают гиперергическими независимо от размера инфильтрата. Гиперемию любого размера без инфильтрата воспринимают как сомнительный результат, справедливо полагая, что в таких случаях не задействованы главные эффекторы ГЗТ — сенсибилизированные Т-лимфоциты и макрофаги.
Гиперчувствительность к туберкулину возникает через 3—8 нед после заражения и сохраняется многие годы, исчезая лишь после элиминации туберкулезной палочки из организма (например, после длительной антибиотикотерапии). Это означает, что по туберкулиновым пробам можно судить о зараженности микобактериями туберкулеза, картируя зоны эндемичности туберкулезной инфекции. Труднее дифференцировать активные и излеченные формы болезни. Наиболее надежны реакции высокой интенсивности, усиление туберкулиновых проб (на 6 мм и более) или переход от отрицательной к положительной реакции (вираж туберкулиновой пробы). Это отражает активацию эндогенной инфекции или реинфицирование извне. Лица с гиперергическими реакциями чаще страдают туберкулезом, а у детей повышение чувствительности к туберкулину нередко предшествует заболеванию. Вообще, туберкулиновые пробы наиболее надежны именно в детском возрасте, когда вероятность неактивных очагов достаточно мала.
Отрицательные пробы у здоровых людей (так называемая положительная анергия) настораживают при риске заражения туберкулезом. Впрочем, негативные тесты не всегда говорят об отсутствии противотуберкулезного иммунитета. Дело в том, что объем, занимаемый агрегатами мононуклеарных клеток вокруг капилляров и придатков кожи (ключевой гистологический признак ГЗТ), напрямую не связан с диаметром уплотнения: выраженная инфильтрация на биопсийных препаратах иногда наблюдается и при отрицательных пробах с туберкулином.
Что касается диагностики непосредственно туберкулеза, то несмотря на то, что чувствительность к туберкулину у больных с активными формами заболевания в среднем выше, чем при латентной инфекции, возможны исключения, которые снижают достоверность результата. Туберкулиновые пробы слабеют и даже становятся отрицательными у больных злокачественными новообразованиями (особенно на фоне лимфогранулематоза), при аутоиммуной патологии, иммунодепрессивной терапии, истощении, острых вирусных инфекциях, СПИДе. Дети, как правило, не реагируют на туберкулин в экзантемном периоде кори, но, выздоравливая, обретают прежний уровень туберкулинопозитивности. Кожная чувствительность к туберкулину исчезает и у больных с тяжелым прогрессирующим течением туберкулеза. Причины этой так называемой негативной анергии не ясны. Они могут быть связаны:
- с избытком гуморальных блокирующих факторов (например, микобактериальных микозидов, острофазных белков);
- задержкой сенсибилизированных Т-лимфоцитов в лимфатических узлах (для туберкулеза характерна лимфопения за счет обратимого дефицита CD4+ Т-клеток);
- гиперпродукцией цитокинов, супрессирующих аллергические реакции.
Подобно другим системным заболеваниям, цитокиновый профиль при туберкулезе варьирует в разных участках тела, отражая степень их вовлечения в патологический процесс. Поэтому анергия кожи не всегда сочетается с ослаблением чувствительности к туберкулину других тканей. Следует, наконец, помнить, что повышенная реактивность к туберкулину возникает после специфической вакцинации. Она обязательна в России, причем первая прививка делается тотчас после рождения. Поствакцинальная аллергия слабее постинфекционной, а главное — быстрее угасает.
В целом пробы с туберкулином сыграли большую роль в изучении туберкулеза и до сих пор, несмотря на ограничения и противоречия, остаются одним из главных эпидемиологических маркеров туберкулезной инфекции, помогая оперативному выявлению лиц, подлежащих усиленному контролю. Есть мнение, что туберкулинодиагностика не имеет диагностического значения у взрослых и может применяться только в педиатрии.
Что касается серодиагностики (т.е. выявления противотуберкулезных антител), то сегодня нет стандартного микобактериального антигена, который можно было бы использовать с этой целью. Предложения ряда авторов не получили признания.
Вакцинопрофилактика
Иммунитет, приобретаемый в результате туберкулезной инфекции, обеспечивает достаточно надежную защиту от повторного заражения и реактивации эндогенного инфекта. Принципиально, что обычно это происходит без клинического конфликта, под влиянием бессимптомной или малосимптомной инфекции. Современная вакцина против туберкулеза представляет собой живые туберкулезные микобактерии бычьего типа, лишенные вирулентности, но сохранившие способность выживать в организме и индуцировать иммунный ответ, похожий на тот, который возникает при естественном заражении вирулентными штаммами.
Вакцина разработана французскими микробиологами А. Кальметтом (Calmette) и Ш. Гереном (Guerin) на основе штамма М. bovis, аттенуированного многочисленными пассажами через среду из картофеля, пропитанного бычьей желчью с 5% глицерина. В честь своих творцов вакцинный штамм получил название бацилл Кальметта—Герена, или BCG (БЦЖ). Такой же акроним закрепился за вакциной — вакцина БЦЖ. Ее широкое внедрение в практику, декретированное Лигой Наций в 1923 г., чуть было не сорвалось из-за трагической случайности, когда в 1930 г. по недосмотру в германском городе Любеке вместо БЦЖ был использован вирулентный штамм М. tuberculosis. Это привело к гибели 73 детей. Пережив шок, БЦЖ-вакцинация (или, как ее называют практики, БЦЖирование) вошла в арсенал стандартных прививок, став опорой в борьбе с туберкулезом. В России БЦЖ-вакцинация обязательна. Ее начинают тотчас после рождения (4—7-й день жизни) и повторяют в возрасте 7 и 13—14 лет.
К сожалению, БЦЖ-вакцина не решила проблему ликвидации туберкулеза. Мировой опыт ее применения зафиксировал противоречивые результаты — от сомнений в отдельных качествах до полного отрицания эффективности. Породив иллюзию безопасности, она скорее содействовала падению интереса к туберкулезу, чем его ликвидации. Так или иначе, но в борьбе с туберкулезной инфекцией современные прививки не могут конкурировать с социальными и санитарными мерами, а лишь дополняют их. Не случайно в большинстве развитых стран БЦЖ-вакцинация отменена или необязательна. Компромиссом является признание того, что, существенно не влияя на показатели инфицирования, вакцинация сокращает число тяжелых, прежде всего внелегочных форм первичного туберкулеза. Поэтому за неимением лучшего БЦЖ-вакцинация признана целесообразной в регионах, неблагополучных по туберкулезной инфекции.
Не прекращаются поиски новых вакцин. В них отражено стремление воспроизвести Т-зависимый противотуберкулезный иммунитет на основе парциальных антигенов и даже дискретных эпитопов микобактерий. Следует, однако, помнить, что протективные факторы БЦЖ до сих пор не идентифицированы, а убитые бактерии БЦЖ лишены защитного эффекта. Такая же участь может постичь субъединичные вакцины, сконструированные из антигенов, вырванных из контекста не только инфекционного процесса, но и адъювантного микроокружения микобактериальных клеток.
Ведутся исследования по созданию вакцины на основе микобактерий-сапрофитов, безвредных для человека. Такой подход основан на изучении возможности воспроизведения перекрестного иммунитета между разными видами микобактерий (в частности, BCG в равной мере защищает от туберкулеза, проказы и условно-патогенных микобактерий). С этой целью испытывается быстрорастущий вид Mycobacterium vaccae. He вызывая обострений у туберкулинпозитивных лиц, такой препарат может оказаться полезным не только для профилактики, но и для иммунотерапии активного туберкулеза.
БЦЖ-вакцина интересует не только фтизиатров. В качестве дешевого, безопасного и иммуноадъювантного субстрата она привлекает внимание специалистов по генноинженерному конструированию вакцинных препаратов. Предпринимаются попытки использовать БЦЖ в качестве носителя генов других бактерий и вирусов, создав таким образом поливалентную вакцину против нескольких инфекций.
Этиотропная терапия
Микобактерии не чувствительны к большинству препаратов, используемых для лечения бактериальных инфекций, что скорее всего связано с плохой проницаемостью их липидной оболочки. Тем не менее известны синтетические и природные антибиотики, которые, убивая или задерживая рост туберкулезных микобактерий, успешно используются в лечении туберкулеза. К числу основных принадлежат изониазид, пиразинамид, рифампицин, стрептомицин, этамбутол. Реже применяют резервные препараты: аминогликозиды (канамицин, капреомицин и др.), циклосерин, этионамид, фторхинолоны, парааминосалициловую кислоту, тиоацетазон.
Терапевтическая стратегия базируется на особенностях туберкулезных поражений. В типичных случаях вторичного туберкулеза они включают:
- каверну, где активно размножаются внеклеточные бактерии;
- некротическую ткань, которая окружает каверну, содержит меньше туберкулезных палочек и имеет кислую рН;
- вал гранулематозной реакции с небольшим числом медленно реплицирующихся (дремлющих) в макрофагах бактерий.
Антибиотики неодинаково активны против каждой из этих субпопуляций возбудителя и поэтому применяются в сочетаниях. Например, изониазид и этамбутол убивают бактерии внутри каверны, быстро сокращая их выделение с мокротой. Вместе с тем они слабо действуют на вяло размножающиеся туберкулезные палочки, не обеспечивая стерилизующего эффекта. Стрептомицин тоже убивает внеклеточные бактерии, но он активен только в щелочной среде и инертен внутри казеозной массы. Пиразинамид действует бактерицидно при низких рН (т.е. в зоне активной воспалительной реакции) и, кроме того, уничтожает внутримакрофагальные бактерии. Рифампицин обладает уникальной способностью убивать дремлющие туберкулезные бактерии, которые периодически проходят через вспышки репликации.
Что касается механизмов действия антитуберкулезных препаратов, то здесь пока не все очевидно. Рифампицин ингибирует активность ДНК-зависимой РНК-полимеразы и, следовательно, синтез белка. Стрептомицин и остальные аминогликозиды тоже подавляют белковый синтез, но на уровне трансляции. Эффект изониазида, этионамида и других агентов связан, по-видимому, с торможением синтеза миколовых кислот. Важно, что препараты разных классов не создают перекрестной резистентности, уничтожая устойчивые друг к другу штаммы.
Это отвечает задаче борьбы с антибиотикорезистентными микобактериями — важным объектом современной фтизиатрии. Сочетание нескольких препаратов необходимо не только для того, чтобы уничтожить различные тканевые субпопуляции бактерий, но и для того, чтобы воспрепятствовать выживанию антибиотикорезистентных клонов, возникающих в результате внутрипопуляционной изменчивости. К сожалению, полностью избежать этого едва ли возможно. В последние годы все более широкое распространение получают штаммы, устойчивые не только к монопрепаратам, но и сразу к нескольким агентам. Они эволюционируют в регионах, где высокая заболеваемость туберкулезом сочетается с неполноценной этиотропной терапией, способствующей отбору и закреплению лекарственноустойчивых мутантов. Лечение должно быть длительным — 9—12 мес. Быстрое прекращение бацилловыделения (иногда через 1—2 мес) обманчиво и не гарантирует полной элиминации возбудителя.
Клостридии
Друзья по жизни мы, враги мы по несчастью.
- Экология: стратегия и тактика.
- Смертельно опасные сапрофиты.
- C. perfringens: лидер гистотоксических клостридий.
- C. perfringens: пищевые инфекции.
- C. difficile: псевдомембранозный антибиотикозависимый колит.
- C. tetani: от раневой инфекции к нейроинтоксикации.
- С. botulinum: нейроинтоксикация без инфекции.
Род клостридий (Clostridium) включает более 80 видов облигатно анаэробных, спорообразующих, крупных (3—10х0,4—1,2 мкм), грамположительных, обычно подвижных (перитрихии), палочковидных бактерий. Споры (точнее эндоспоры) вызывают утолщение клеток, придавая им вид веретена (рис. 1). Отсюда и название — клостридии (греч. kloster — веретено). Неоднородность клостридий проявляется в морфологии спор и вегетативных клеток, в чувствительности к кислороду, устойчивости во внешней среде, в условиях спорообразования и метаболической активности. Являясь сапрофитами, освобождающими природу от мертвого органического материала (через процессы брожения и гниения), клостридии обладают неодинаковой болезнетворностью. Лишь немногие из них вступают в прижизненные контакты с человеком и животными, обладая условной патогенностью. Присутствие болезнетворных клостридий в кишечнике (в том числе человека) не может поколебать отношения к ним как к сапрофитам: здесь они тоже питаются органическими остатками, не инвазируя живой ткани. Следует скорее говорить о том, что клостридии охотно меняют независимый образ жизни на симбиоз, перекладывая на хозяина заботы о взаимоотношениях с внешней средой. Оказавшись на свободе, они не только выживают в виде спор, дожидаясь очередной вегетации в организме хозяина, но и активно размножаются.

Являясь сапрофитами, освобождающими природу от мертвого органического материала (через процессы брожения и гниения), клостридии обладают неодинаковой болезнетворностью. Лишь немногие из них вступают в прижизненные контакты с человеком и животными, обладая условной патогенностью. Присутствие болезнетворных клостридий в кишечнике (в том числе человека) не может поколебать отношения к ним как к сапрофитам: здесь они тоже питаются органическими остатками, не инвазируя живой ткани. Следует скорее говорить о том, что клостридии охотно меняют независимый образ жизни на симбиоз, перекладывая на хозяина заботы о взаимоотношениях с внешней средой. Оказавшись на свободе, они не только выживают в виде спор, дожидаясь очередной вегетации в организме хозяина, но и активно размножаются.
Клостридии вызывают три смертельно опасных заболевания — газовую гангрену (C. perfringens и другие гистотоксические клостридии), столбняк (C. tetani) и ботулизм (C. botulinum). Кроме того, они причастны к патологии пищеварительного тракта, включая относительно доброкачественные энтероколиты и деструктивные процессы, требующие активного врачевания (С. perfringens, C. difficile). Патогенность связана со способностью продуцировать мощные токсины, которые образуются в инфицированном организме или во внешней среде (C. botulinum). Инвазивность почти всегда ограничена, и даже при массивных поражениях (газовая гангрена) носит вторичный характер, следуя за анаэробным повреждением тканей. Не случайно о клостридиях говорят как о некропаразитах, подчеркивая их неспособность к размножению в живых, хорошо снабжаемых кровью тканях.
Clostridium perfringens
Подобно другим клостридиям, C. perfringens (син. С. welchii) широко распространены во внешней среде, особенно в унавоженных почвах. Они сохраняются здесь не только в виде спор, но и активно вегетируют, особенно при повышенной (45°С) температуре. Вместе с тем представители этого вида, пожалуй, больше, чем другие клостридии, экологически связаны с кишечником человека и животных. У человека они определяются в количестве 103—105 бактерий на 1 г фекалий (для сравнения: содержание факультативно-анаэробных бактерий, прежде всего кишечной палочки и энтерококков, составляет 106—107/г, а бактероидов и бифидобактерий, доминирующих в кишечном микробиоценозе, — 109—1010/г).
Видовой эпитет (perfringens) означает «прорывающий», «разламывающий». Он отражает мощную ферментативную активность данного вида клостридий, которые благодаря энергичному газообразованию разрывают питательные среды и столь же активно ведут себя в инфицированных тканях (рис. 2).


По спектру продуцируемых токсинов различают пять типов C. perfringens — A, B, C, D, E. Заболевания человека чаще связаны с типом А (газовая гангрена, пищевые инфекции) и изредка — С (некротизирующий энтероколит). В целом понятие «токсин» у C. perfringens довольно расплывчато. Оно объединяет не менее 14 факторов с летальной и гистолитической активностью. Главными являются альфа-, бета, эпсилон- и йота-токсины. Именно их комбинация определяет тип C.perfringens. Альфа-токсин продуцируют только клостридии типа А. Это мощный цитотоксин, обладающий свойствами фосфолипазы С. Он расщепляет фосфолипиды (лецитин) клеточных мембран, лизируя клетки (рис. 3).

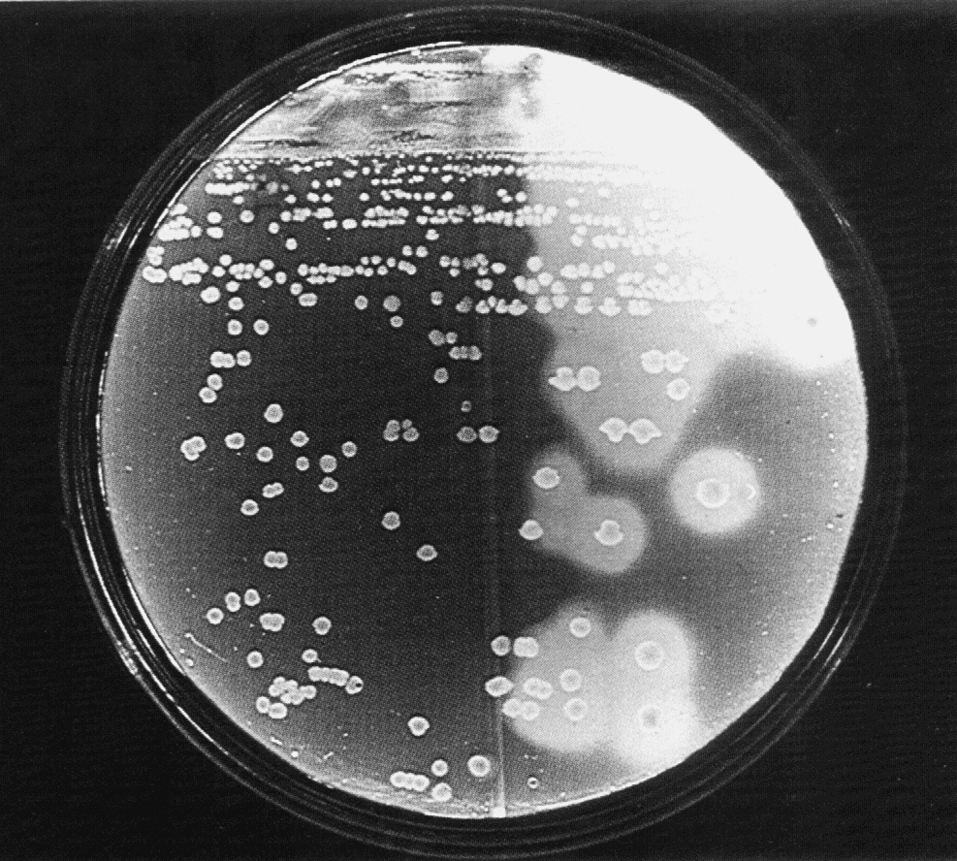
При газовой гангрене альфа-токсин служит не только главным эффектором внутритканевой инвазии, но и причиной внутрисосудистого гемолиза — символа смертельной клостридиальной токсинемии. Из других (минорных) токсинов C. perfringens типа А образует тета-токсин, или перфринголизин (чувствительный к кислороду гемолизин, похожий на О-стрептолизин стрептококка), каппа-токсин (коллагеназа), мю-токсин (гиалуронидаза) и ню-токсин (ДНК-аза). Они лишены самостоятельного значения, но усиливают действие альфа-токсина.
Штаммы типа А, вызывающие пищевое отравление, продуцируют энтеротоксин. Это термолабильный белок с молекулярной массой 35 кДа, который высвобождается в тонком кишечнике при споруляции (образовании спор). Подобно энтеротоксинам холерного вибриона и энтеробактерий, он усиливает секрецию энтероцитами Na+, Cl— и воды, но достигает этого иным путем: он не повышает внутриклеточного уровня цАМФ, а повреждает плазматическую мембрану энтероцитов. Это единственный такого рода пример среди диареегенных бактерий. Особняком стоит энтеропатогенность C. perfringens типа С. Она связана с бета-токсином и проявляется в виде некротизирующего энтерита (см. ниже).
Газовая гангрена. Гангреной (греч. gangraina — разъедающая язва) называется область омертвения (некроза) тканей, которая подвергается гнилостному распаду под влиянием бактерий. Внедрению последних способствует то, что некротизированная ткань не снабжается кровью, лишаясь притока антибактериальных факторов и способности к воспалительной реакции. В сочетании с анаэробиозом это создает идеальную среду для размножения и реализации болезнетворного потенциала анаэробных сапрофитов. Не обладая достаточной агрессивностью для инвазии живой ткани, они играют главную роль в гангренозных поражениях.
Газовая гангрена — эталон тяжелейших последствий, связанных с клостридиальным заражением некротизированных тканей. Процесс эволюционирует по типу мионекроза — скоротечного поражения мышц с ничтожными признаками воспаления и обильной продукцией газа. К этому предрасполагает размозжение мышечной ткани, которое вызывает местную аноксию и открывает путь для бактериальных спор и вегетативных клеток. Они попадают сюда из внешней среды или из собственной микрофлоры, обычно с инородными телами (почва, обрывки одежды, осколки снарядов и пр.). Чаще всего это наблюдается при боевых ранениях («болезнь войн»), а в мирное время — при травмах, полученных в автомобильных и иных катастрофах, после оперативных вмешательств (особенно лапаротомии) и даже обычных инъекций, при кишечной непроходимости и пролежнях. В целом это оправдывает определение газовой гангрены как «раневой инфекции».
Вегетации клостридий способствует коинфицирование аэробными и факультативно анаэробными бактериями: редуцируя кислород, они усиливают анаэробиоз. Преобладание C. perfringens объясняется несколькими причинами: 1) они чаще других клостридий представлены в почве и микрофлоре человека; 2) обладают одним из самых мощных некротизирующих токсинов — альфа-токсином; 3) относительно аэротолерантны, а потому менее требовательны к ишемизации тканей; 4) обладают капсулой, усиливающей инвазивность (рис. 4).

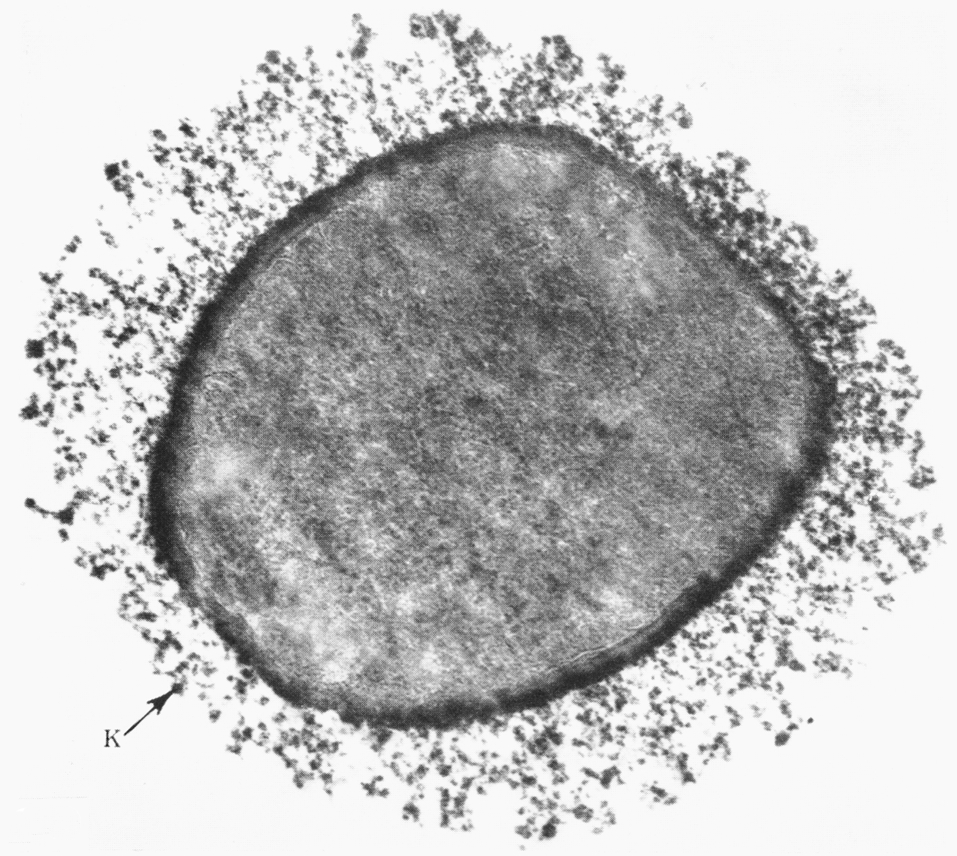
После стабилизации инфекта в зоне первичного (травматического или ишемического) некроза клостридии размножаются с выделением альфа-токсина и ферментов, которые расширяют зону некроза, убивая здоровые клетки соседней мышечной ткани (альфа-токсин) и разрушая соединительнотканный матрикс (коллагеназа и др.). Ферментация миоцитарного гликогена ведет к накоплению молочной кислоты и газа, в основном двуокиси углерода и водорода. Газообразование определяет характерный симптом клостридиального мионекроза — крепитацию (лат. crepitare — скрипеть, хрустеть), а кислая среда активирует протеолитические ферменты мышечной ткани и аутолитическое высвобождение аминокислот — великолепного питания для клостридий. Некротические изменения сосудов вызывают отек (отсюда одно из старых названий болезни — злокачественный отек), что вкупе с накоплением газа повышает внутритканевое давление, углубляя ишемию и тканевой анаэробиоз. Зоны поражения бедны нейтрофилами, которые либо не доходят сюда из-за прекращения кровотока, либо разрушаются клостридиальными токсинами. На фоне быстро прогрессирующего мионекроза бактерии захватывают новые участки тканей, распространяясь вслед за деструктивными токсинами и ферментами. Для этого им не требуется активной подвижности: в отличие от остальных клостридий C. perfringens лишены жгутиков. Кожа чернеет, лопаясь под давлением жидкости и газа (рис. 5). Всасываясь в кровь, бактериальные токсины (прежде всего альфа-токсин) и продукты тканевого распада вызывают гемолиз, гемоглобинурию и почечную недостаточность. Бактериемия не имеет патогенетического значения и наблюдается лишь в терминальной фазе. Смерть наступает от гипотонии периферических сосудов (шок).

Кроме C. perfringens (выделяется от 60% больных), причиной заболевания могут быть и другие клостридии — C. oedematiens (C. novyi), C. septicum (20—40%), реже C. histolуticum. В целом бактериология газовой гангрены варьирует. Часто это смешанная инфекция, так как контаминирующий материал содержит множество бактерий, включая аэробные (синегнойная палочка) и факультативно анаэробные (стрептококки, стафилококки, колиформные бактерии) виды. Они не вызывают некроза мышечной ткани, но помогают анаэробам, редуцируя кислород.
Клостридии вовлекаются и в менее тяжелые раневые процессы типа флегмоны, или целлюлита. В этом случае поражается подкожная клетчатка, мышцы не затрагиваются. Если бактерии не находят условий для размножения, контаминация не имеет клинических последствий.
Патогенетическое лидерство клостридиальных токсинов при газовой гангрене стимулировало множество исследований по специфической профилактике и лечению раневой инфекции. Были созданы анатоксины и антитоксические сыворотки, широко применявшиеся в годы войны. Однако их эффект скорее иллюзорен, и сегодня они не применяются. Это сопряжено с особенностями патогенеза газовой гангрены: бактерии размножаются и выделяют токсины в тканях, лишенных кровоснабжения (антитела сюда не доходят), а общая интоксикация носит сложный характер, включая действие продуктов тканевого распада. При помощи антитоксинов удается предупредить развитие клостридиальной инфекции у животных, но они не обрывают реального процесса.
Клостридии чувствительны к различным антибиотикам (в частности, к пенициллину), но они, не достигая мишени (отсутствие кровотока!), помогают мало. Наиболее надежна хирургическая обработка ран, нацеленная на ликвидацию анаэробиоза (удаление мертвых тканей и дренирование ран).
Пищевые инфекции. В 1945 г. был описан острый диарейный синдром после приема пищи, обсемененной C. perfringens. Вслед за стафилококковой интоксикацией и сальмонеллезной инфекцией это самое распространенное пищевое отравление микробной природы. Большинство случаев связано с энтеротоксигенными штаммами типа А, которые отличаются слабой продукцией альфа-токсина, но образуют термолабильный токсин, повреждающий энтероциты (см. выше). Для заболевания характерны схваткообразные боли в животе, зловонный пенящийся понос, изредка рвота и температура. Симптомы возникают через 7—24 ч после приема инфицированной пищи, выражены обычно умеренно и проходят без лечения через 2—3 дня. Для развития заболевания требуется примерно 108 клеток C.perfringens, которые, подвергаясь споруляции в тонком кишечнике, высвобождают энтеротоксин, накопившийся в бактериях при их вегетации в продуктах питания; в пищеварительном канале возбудитель не размножается. Особенно опасна подогретая пища, оставленная на несколько часов при комнатной температуре, так как, выдерживая кипячение в течение часа, споры быстро прорастают при 40—50°С. Вероятность патогенетически значимой контаминации пищевых (как правило, мясных) продуктов повышается благодаря широкому распространению клостридиальных спор.
Тяжелой (к счастью, редкой) формой клостридиальной патологии является некротизирующий энтерит, вызываемый обычно C. perfringens типа С. Характерен кровавый понос, причиной которого является повреждение слизистой оболочки бета-токсином, образующимся при размножении клостридий в тонком кишечнике. Спорадические случаи описаны во многих странах. Особенно показательны вспышки среди аборигенов Новой Гвинеи во время ритуальных пиршеств с обильным поеданием плохо обработанной свинины. Смертность достигает 30—40%.
Clostridium difficile
С. difficile (название возникло из-за трудностей культивирования) открыт в 1935 г., т.е. значительно позже других клостридий. Вслед за почти полным забвением он был признан главным (возможно, единственным) возбудителем псевдомембранозного колита, ассоциированного с применением антибиотиков (клиндамицин, цефалоспорины, ампициллин). Классическим симптомом является госпитальная диарея с примесью крови, которой сопутствуют схваткообразные боли в животе, лейкоцитоз и лихорадка. Очаги поражения, выявляемые в толстом кишечнике, содержат отложения фибрина, остатки некротизированной слизистой оболочки и лейкоцитов (псевдомембрана). Часто процесс протекает в стертых формах, но возможно и фульминантное развитие событий, опасное для жизни.
Подобно всем клостридиям, C. difficile убиквитарен, персистируя в почве, воде и кишечном содержимом многих животных. Вместе с тем возбудитель редко (всего в 3% случаев) выделяется из фекалий здоровых взрослых людей. Носительство возрастает у госпитальных больных (13—30%), отражая обмен микрофлорой при плотных контактах. Отсюда возникает комплекс мероприятий, направленных на предупреждение заражения, особенно в больницах, где уже отмечались вспышки инфекции. Парадоксом является широкая контаминация здоровых детей — 14—50%; у новорожденных она достигает даже 90%. Полагают, что токсины C. difficile не рецептируются энтероцитами новорожденных и не вызывают их поражения.
События, ведущие к диарейному синдрому, начинаются с антибиотикозависимого подавления нормальной микрофлоры, прежде всего анаэробных неспоровых бактерий (бактероидов, бифидобактерий), — основы колонизационной резистентности толстого кишечника. Наиболее опасны антибиотики типа клиндамицина, которые, обладая высокой активностью против анаэробных бактерий, длительно (до 5 дней) сохраняются в кишечнике после однократного приема. C. difficile выживают благодаря образованию спор. Прорастая после отмены антибиотиков, они дают начало обильному потомству вегетативных клеток, продуцирующих токсины.
C. difficile образует два главных токсина — А и В. Они секретируются большинством штаммов, обладая неодинаковым спектром биологической активности, закодированной вместе с позитивным и негативным регулятором в острове патогенности (см. «Болезнетворность бактерий»). Это наиболее крупные (308 и 269 кДа) из однонитевых бактериальных экзотоксинов, которые имеют до 45% сходства в последовательности аминокислот. Рецепторы известны только для токсина А.
В состав токсинов А и В входит каталитический домен, определяющий глюкозилирование G-белков — Rho, Rac и Cdc42. Это лишает G-белки ферментативной способности к трансформации ГДФ—ГТФ, от которой зависят многие важные функции клеток, в том числе полимеризация-деполимеризация актина. Токсин А действует как энтеротоксин. Повреждая мукозальные клетки, он вызывает образование вязкой кровянистой жидкости в просвете изолированной кишки кролика. Агрессивность токсина В более универсальна. В культуре клеток он в 1000 раз токсичнее, чем токсин А, приводя к округлению клеток благодаря повреждению цитоскелета. Антитоксический иммунитет, который удается воспроизвести на животных, клинически не надежен, так как токсины быстро связываются чувствительными клетками, ускользая от нейтрализующих антител. Выявление токсинов в фекалиях рекомендовано для подтверждения клинического диагноза. Это много проще, чем выделение и идентификация культуры C. difficile.
Лечение нацелено на уничтожение вегетирующих клеток C. difficile. Эффективны метронидазол и ванкомицин, которые благодаря плохой всасываемости при пероральном приеме накапливаются в толстом кишечнике. Проблемой остаются рецидивы, поскольку антибиотики не гарантируют полного освобождения кишечника от спор и даже способствуют их образованию. В связи с этим логичным выглядит восстановление нормального микробиоценоза при помощи пробиотиков, т.е. живых нормальных бактерий толстого кишечника. Положительный опыт такого рода экспериментально обоснован, но клиническая значимость современных препаратов (на основе бифидобактерий, лактобацилл, кишечной палочки) сомнительна. Определенные надежды связывают (и уже пытаются реализовать) с дрожжами Saccharomyces boulardii, которые не очень понятным образом тормозят размножение C. difficile и образование их токсинов в кишечнике. Есть данные, говорящие о том, что S. boulardii выделяют протеазу, разрушающую клостридиальные токсины, или действуют как фактор, блокирующий транскрипционные процессы в эпителиоцитах, необходимые для передачи токсигенных сигналов С. difficile.
Clostridium tetani
Первые описания столбняка (tetanus) встречаются у Гиппократа и других врачей древности. Инфекционная природа заболевания экспериментально обоснована Карлэ и Раттом в 1884 г. Остроумные опыты (1884—1886 гг.) провел Николайер. Он показал, что инокуляция взвеси почвы вызывает столбняк у животных. Обнаружив в зоне инъекции длинные, тонкие палочки, отсутствовавшие в других тканях, Николайер предположил, что возбудитель размножается местно, продуцируя стрихниноподобный яд — непосредственную причину болезни. Розенбах (1886—1887 гг.) подтвердил наблюдения Николайера, выявив у столбнячных больных спорообразующие палочки. Круг сомкнулся, когда Китазато (1889 г.) воспроизвел столбняк при введении животным чистой культуры возбудителя. Перед анаэробным культивированием раневого содержимого от больного столбняком он догадался уничтожить неспоровые бактерии, подвергнув исследуемый субстрат термической обработке.
Характерным морфологическим признаком столбнячных бацилл являются круглые терминальные споры, благодаря которым они выглядят как булавки или барабанные палочки (рис. 6).

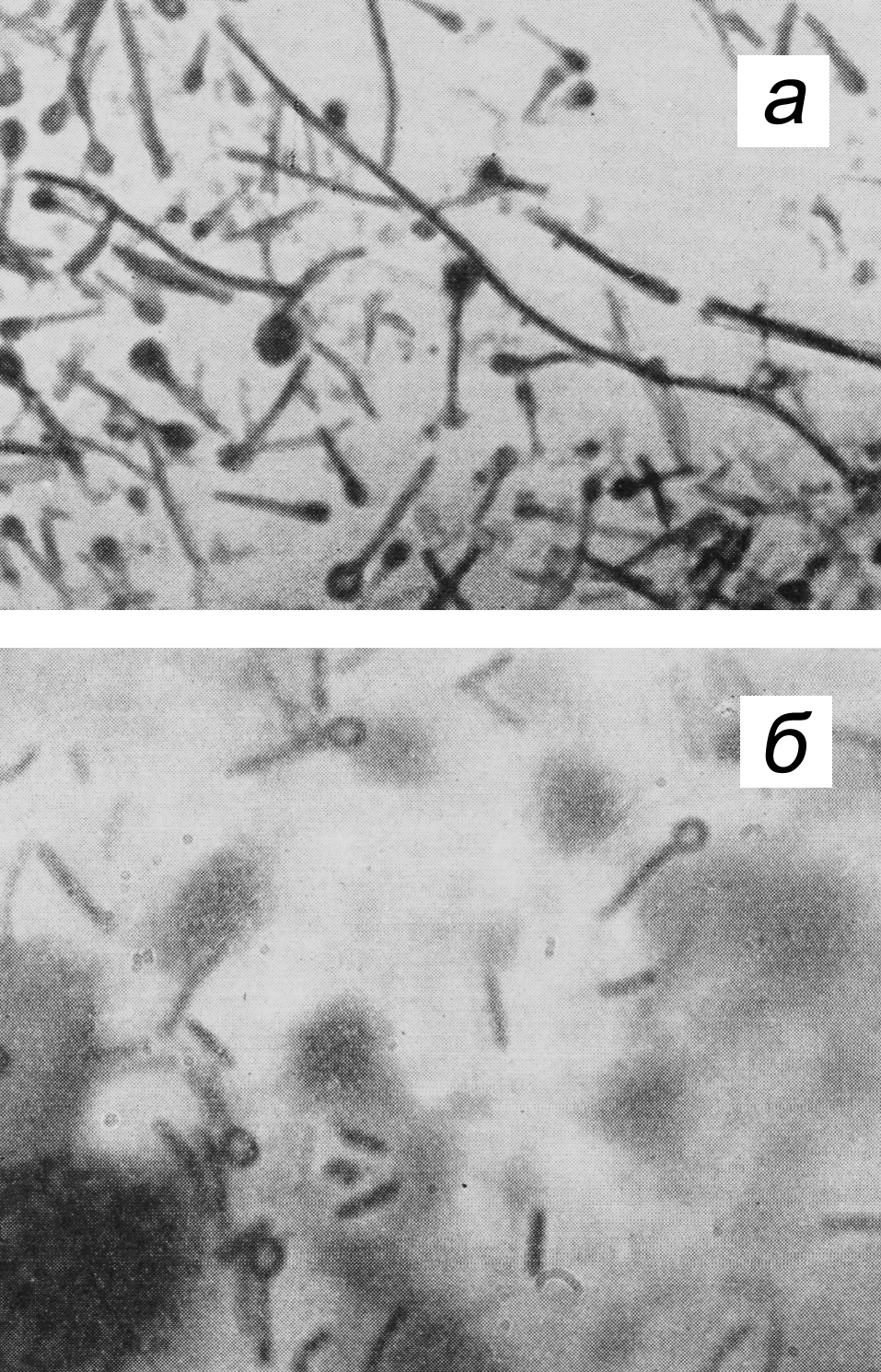
В культурах споры образуются через несколько суток и, по разным данным, переносят кипячение от 20 мин до 3 ч. Столбнячные палочки подвижны и по антигенным особенностям жгутиков подразделяются по меньшей мере на десять серотипов. Впрочем, это не имеет практического значения, так как все разновидности C. tetani вырабатывают идентичный нейротоксин, что обеспечивает единство патогенеза и унификацию иммунопрофилактики.
Клостридии столбняка широко распространены в природе, прежде всего в почве, особенно на возделываемых (окультуренных) участках. Нередко они обнаруживаются и в кишечнике человека и животных, хотя стиль их жизни в этом случае остается загадкой. Возможно, столбнячные бациллы здесь не размножаются, проходя транзитом в виде спор. Не исключена и вегетация, о чем говорят следы противостолбнячного антитоксина в сыворотке здоровых животных. Но если это и так, столбняк никогда не возникает при всасывании токсина из кишечника, тем более что вызвать заболевание путем перорального введения токсина не удается.
Столбняк развивается как раневая инфекция, но далеко не каждое ранение создает условия для прорастания спор и размножения бактерий. C. tetani принадлежит к числу наиболее строгих анаэробов, и во многих случаях это спасает от фатальных последствий. Глубокая гипоксия, необходимая для токсинообразования, как правило, возникает в зоне некроза, и все, что было сказано о возбудителях газовой гангрены, справедливо для cтолбнячных клостридий. Однако, отмечая опасность тяжелых травм, следует помнить, что даже малые повреждения могут стать причиной столбняка. Коварны колотые раны (от гвоздей, заноз, колючек и пр.), в глубине которых возникает анаэробиоз. Не случайно у 20% больных отсутствуют указания на травму. Это не означает, что ее не было. Просто она не запомнилась из-за незначительности. Есть подозрения, что столбнячные споры могут персистировать в тканях. По крайней мере известны случаи столбняка, спровоцированные локальной ишемией спустя более 10 лет после предполагаемого заражения. У новорожденных инфекция проникает через пупочный канатик, так что tetanus neonatarum (рис. 7) остается реальностью в странах с низкой санитарной культурой и плохо налаженной иммунопрофилактикой. Беременные женщины должны вакцинироваться столбнячным анатоксином, чтобы обеспечить пассивный иммунитет новорожденных.

В отличие от гистотоксических клостридий, C. tetani лишен инвазивности (возбудитель не покидает места первичного заражения), и заболевание всегда протекает как интоксикация на фоне малосимптомной очаговой инфекции. Столбнячный нейротоксин называется тетаноспазмином, что подчеркивает ведущий, по сути единственный патогенетический механизм столбняка — спастические параличи поперечно-полосатых мышц. Столбнячный токсин принадлежит к классике микробной токсинологии, уступая по биологической активности только ботулиническому яду. Вслед за человеком к нему наиболее восприимчивы лошади, белые мыши и морские свинки. Птицы и хладнокровные животные резистентны к столбняку. Смертельная доза для мыши составляет всего 3·10-11 г, т.е. 1 мг достаточно, чтобы убить 50—70 млн. мышей.
Способность к токсинообразованию закодирована в плазмиде и неодинаково выражена у разных штаммов — от нетоксигенных до высокотоксигенных. Токсин накапливается внутри клеток и высвобождается при аутолизе и образовании спор. Это белок с молекулярной массой 150 кДа, который состоит из двух цепей (тяжелой и легкой) и активируется собственными (C. tetani) и тканевыми протеазами. Этапность интоксикации сводится к следующему. Тяжелая цепь активированного токсина рецептируется нервными окончаниями в нервно-мышечных синапсах благодаря связыванию с ганглиозидами пресинаптической мембраны мотонейронов. Рецепция отличается высокой избирательностью, объясняя поразительную токсичность клостридиальных нейротоксинов, прицельно атакующих нейроны. После эндоцитоза нервными окончаниями комплекс «рецептор—токсин» вливается в поток ретроградного внутриаксонального транспорта, достигая передних рогов спинного мозга. Здесь, после высвобождения в синаптическое пространство между телом мотонейрона и тормозящими интернейронами, токсин вновь подвергается эндоцитозу — на этот раз тормозящими нейронами. В дело вступает легкая субъединица. Обладая активностью цинкзависимой эндопептидазы, она инактивирует факторы (синаптобревины), необходимые для секреции медиаторов (глицина, гамма-аминобутировой кислоты), блокирующих возбуждение мотонейронов. Это повышает их раздражимость и ведет к стойким мышечным контрактурам — спастическим параличам. При достаточной дозировке токсина в процесс вовлекаются соседние мотонейроны, что вызывает генерализацию мышечных контрактур.
Развитие симптомов при введении токсина мыши под кожу задней конечности выглядит следующим образом. Сначала обнаруживаются признаки местного столбняка: нога вытягивается, как палка, и при движениях беспомощно тащится, хвост становится ригидным и изогнутым. Спазм распространяется на мышцы той же половины туловища, приводя к искривлению тела в сторону пораженной конечности (рис. 8). Контрактуры захватывают противоположную сторону тела, и животное гибнет от общего столбняка.


Такая картина типична для мелких животных и называется восходящим столбняком (tetanus ascendens). У человека последовательность мышечных поражений не связана с зоной первичного токсинообразования, а имеет характер нисходящего процесса — tetanus descendens. Он начинается со сведения жевательных мышц (тризм; греч. trismos — стискивание, сжатие) и затылочной мускулатуры. Затем спазмируются мышцы лица, обретающего подобие вынужденной улыбки (risus sardonicus; см. рис. 7), и далее — мускулатура туловища и конечностей. Поскольку мышцы спины сильнее мышц живота, тело напоминает дугу — опистотонус (греч. opisthios — задний, tonos — напряжение; рис. 9).

Из-за повышенной возбудимости мотонейронов к мышечным спазмам присоединяются приступы судорог даже от слабых раздражителей (шум, движение воздуха). Чувствительная сфера не затрагивается, больной остается в сознании. Смерть наступает от асфиксии из-за паралича мышц грудной клетки и диафрагмы.
Условия для нисходящего столбняка возникают после всасывания токсина в кровь (токсинемия). С кровью он разносится по тканям, где фиксируется окончаниями двигательных нервов. Восходящий столбняк протекает без токсинемии, в результате связывания токсина нервными окончаниями в зоне образования или введения. Внутрисосудистая инъекция токсина всегда вызывает нисходящий столбняк. При этом этапность спастического поражения мышц (в частности, опережающее вовлечение мышц головы и шеи) отражает их удаленность от центральной нервной системы, т.е. расстояние, которое должен пройти токсин до своих мишеней — мотонейронов, иннервирующих соответствующую группу мышц (средняя скорость внутриаксонального тока жидкости — 250 мм в сутки). Этим в известной мере определяется и длительность инкубационного периода: у человека он составляет 7—10 дней (с колебаниями от 2 до 50 дней), а у животных возрастает с увеличением размеров тела (например, 12 ч у мыши и 5 дней у лошади).
Даже при лечении антитоксической сывороткой смертность составляет 15—80% и в целом обратно пропорциональна продолжительности инкубации. Критическими являются первые дни заболевания, отражая старинное правило Гиппократа: «Люди, схваченные столбняком, умирают в течение четырех дней или не умирают вообще». Столбняк не оставляет иммунитета, так как ничтожное количество токсина, продуцируемое возбудителем, быстро связывается нервной тканью, ускользая от иммунной реакции. Вместе с тем токсин, обезвреженный формалином (анатоксин), создает великолепный иммунитет, и при хорошо отлаженной иммунопрофилактике столбняк — большая редкость. Экстренную профилактику проводят при помощи антитоксических сывороток или гомологичного (человеческого) противостолбнячного иммуноглобулина. При этом надлежит помнить, что время решает все, так как после эндоцитоза токсин не доступен для антител. Неоценимую роль играет хирургическая обработка ран. Показаны и антибиотики, которые полезны не только для подавления столбнячной палочки, но и сопутствующей (кислородредуцирующей) флоры. Подобно другим клостридиям, С. tetani чувствителен к пенициллину, цефалоспорину, тетрациклину, метронидазолу и др.
Для диагноза достаточно характерной клиники. Бактериологического подтверждения обычно не требуется, тем более, что в организме споры образуются не всегда и типичные бациллы (барабанные палочки) выявляются редко. Возбудитель может быть выделен из раны, но работа с анаэробами требует специальных условий, которых лишены стандартные лаборатории. Возможна и биологическая проба на мышах с кровью больного. Но ее результаты непостоянны и воспроизводимы лишь при гипертоксинемии.
Сlostridium botulinum
Ботулинические клостридии вызывают тяжелое пищевое отравление, изначально описанное после употребления зараженной колбасы (лат. botulus — колбаса). Возбудитель был выделен в 1896 г. в Бельгии ван Эрменгемом из ветчины, послужившей причиной группового заболевания с несколькими смертельными исходами. Фильтраты культур, выращенных в строго анаэробных условиях, содержали токсин и (так же, как экстракт ветчины) вызывали характерные симптомы при скармливании животным (кошкам).
Для C. botulinum характерны споры овальной формы, которые располагаются субтерминально и придают палочкам вид теннисной ракетки (см. рис. 1). Они принадлежат к числу наиболее устойчивых спор, выдерживая кипячение в течение нескольких (по некоторым данным до 20) часов. Автоклавирование при 120°С убивает их через 20 мин. Токсин ботулинических клостридий тоже относительно стабилен: при 80°С он разрушается через 30—40 мин, а при 100°С — через 10 мин.
С практической точки зрения ботулинические клостридии удобно рассматривать как единый вид, хотя на самом деле это довольно разнородная группа бактерий, продуцирующих фармакологически сходный, но антигенно различный токсин. Известно 8 серологических вариантов ботулинического токсина и, соответственно, 8 серотипов C. botulinum — A, B, C1, C2, D, E, F, G. Токсины С, D и Е закодированы в умеренных фагах, остальные — в хромосоме.
Ботулинический токсин считается самым сильным ядом биологического происхождения, но по неясным причинам чувствительность разных животных к его вариантам неодинакова. Человек поражается только токсинами А, В и Е. Непонятно, почему некоторые животные (например, крысы, собаки, куры) отлично переносят ботулинический яд, а другие (мыши, морские свинки, кошки, лошади, утки) высокочувствительны к нему. Человек, по-видимому, наиболее восприимчив к ботулизму. Особенно опасен токсин А, смертельная доза которого для человека составляет всего 10-8 г (6 кг достаточно, чтобы отравить все человечество!).
Из всех патогенных клостридий ботулинические клостридии больше всего отвечают критериям сапрофитизма. Они широко распространены не только в окультуренной почве, но и в девственных регионах, не связанных с жизнедеятельностью человека. Их присутствие доказано в иле рек и озер, откуда они попадают в кишечник рыб и водоплавающих птиц. Реже, чем другие клостридии, C. botulinum обнаруживается в пищеварительном канале домашних животных и лишь изредка у человека. Многие штаммы лучше размножаются при 25°С, что также подчеркивает независимость от теплокровных животных.
Благодаря широкому распространению споры C. botulinum могут инфицировать любые продукты растительного и животного происхождения. Вместе с тем ботулизм — редкое заболевание, так как возбудитель обычно не размножается в организме или по крайней мере не продуцирует в нем токсина. Поэтому свежая (сырая или вскоре после кулинарной обработки) пища не представляет опасности. Угроза возникает лишь при прорастании спор и размножении C. botulinum. В подходящих условиях для накопления патогенетически значимой концентрации токсина в культуре клостридий типа А достаточно суток. Хранение в холодильнике не снимает опасности, так как токсин (особенно типа Е) медленно (через несколько недель) образуется даже при 3—5°С. Коварность ботулинических клостридий состоит и в том, что продукция токсина почти не влияет на вкусовые качества пищи. Можно лишь отметить запах прогорклого масла, который характерен и для чистых культур C. botulinum.
Вегетации бактерий и образованию токсина противодействуют многие факторы, что резко снижает вероятность ботулизма даже при обильном обсеменении продуктов. Так, прорастание спор подавляется в кислой (рН=4,5) среде (соответствует 2% раствору уксуса в маринадах) и при 10% концентрации хлористого натрия (полезно помнить при солении рыбы). Абсолютным условием для токсинообразования служит отсутствие кислорода (закатанные банки с консервами, герметизация в полиэтиленовых пакетах, размножение внутри мясного фарша и пр.). С неравномерностью накопления токсина в продукте и более медленным повышением температуры внутри его массы при термической обработке связаны случаи избирательного поражения ботулизмом после употребления общего продукта.
Токсин высвобождается при аутолизе бактерий (усиливается при образовании спор) в виде комплекса с инертными белками. Они экранируют токсический компонент от протеолитических ферментов и кислой среды желудка. Но эта защита относительна, и при пероральном введении до цели доходит лишь малая часть токсина. Этим объясняется то, что при скармливании токсина чувствительность к нему у животных в 1000—1500 раз меньше, чем при парентеральном введении. По ряду структурно-функциональных признаков ботулинический токсин похож на столбнячный тетаноспазмин. Это полипептидная цепь (молекулярная масса 150 кДа) с одной или несколькими внутримолекулярными дисульфидными связями. В нативном виде малотоксичен, но активность возрастает под влиянием трипсина, тканевых и собственных (клостридиальных) протеиназ. Они расщепляют молекулу на два субкомпонента (легкий и тяжелый), которые до внедрения в чувствительные клетки (нейроны) скреплены дисульфидной связью и соответствуют А- и В-фрагментам бинарных токсинов (см. «Болезнетворность бактерий»).
После всасывания из тонкого кишечника тяжелая субъединица токсина избирательно связывается ганглиозидами пресинаптической мембраны мотонейронов, вызывая эндоцитоз всей молекулы нервными окончаниями. В отличие от столбнячного токсина ее путь на этом заканчивается, т.е. эффект ограничивается периферической нервной системой. После восстановления дисульфидной связи легкая субъединица высвобождается из эндосом и, действуя как эндопептидаза, разрушает факторы (синаптобревины), необходимые для секреции ацетилхолина. Это обрывает нервные импульсы, идущие от мотонейронов к мышечным волокнам, приводя к необратимой релаксации последних — вялые параличи. Вовлечение краниальных нервов ведет к диплопии (двойное видение), дисфагии (затруднение глотания) и дисфонии (затруднение речи). Зрачки расширены и неподвижны. Из-за ослабления или даже прекращения секреции слюны возникает сухость во рту, падает тонус мышц шеи и туловища. Летальность достигает 20—40% на фоне паралича дыхательных мышц или сердечной недостаточности. Сознание сохраняется вплоть до смерти.
Следуя патогенетической логике, больным вводят антитоксические сыворотки против токсинов (А, В и Е), доминирующих при ботулизме у человека. Но, обладая профилактическим эффектом, они приносят мало пользы в борьбе с уже развившимся заболеванием. Как и при столбняке, токсин, транслоцированный в нейроны, недоступен для антител.
Для подтверждения диагноза проводят определение токсина в сыворотке, фекалиях, желудочном содержимом, рвотных массах и экстрактах из пищевых продуктов (биологическая проба на мышах, иммунохимические тесты).
Раневой ботулизм. Клостридии ботулизма способны размножаться и продуцировать токсин при заражении ран, особенно в условиях смешанной инфекции. Это казуистика, но она экологически сближает C. botulinum с другими болезнетворными клостридиями.
Ботулизм младенцев. В качестве самостоятельной нозологической формы утвержден в 1976 г. Поражаются дети в возрасте до 6 мес, в среднем — 2,5 мес. Болезнь обычно протекает легко и не диагностируется. Клинические признаки включают запор (первый симптом), вялость, затруднение сосания и глотания, измененный плач, мышечную слабость. В тяжелых случаях возникают симптомы взрослого ботулизма, но летальность не превышает 4%. Допускают, что ботулиническая интоксикация является причиной некоторых случаев внезапной смерти новорожденных (смерть в колыбели).
Принципиально, что в отличие от ботулизма взрослых младенческий ботулизм развивается как инфекционный процесс: токсин образуется при вегетации спор, попавших в толстый кишечник. Это может быть связано с особенностями кишечной микрофлоры у малышей, не оказывающей должного сопротивления C. botulinum. Источники инфекции остаются загадкой, хотя, памятуя о широком распространении клостридиальных спор, можно назвать массу возможностей для контаминации.
Неклостридиальная анаэробная инфекция (семейство Bacteroidaceae)
От домашнего вора не убережешься.
- Изобилие необъятного.
- Бактероиды и их псевдородственники.
- Отношение к нормальной микрофлоре.
- Патогенность и патология.
- Факторы и реализация болезнетворности.
- Диагностика.
- Чувствительность к антимикробным препаратам.
К семейству Bacteroidaceae относятся грамотрицательные, обычно неподвижные, плеоморфные палочки, иногда веретенообразной формы. Они не образуют спор. Бактероиды — облигатные анаэробы. Чувствительность к кислороду неодинакова у разных видов, но многие достаточно аэротолерантны и выживают (но не размножаются) в обычной атмосфере до 72 ч. Растут на обогащенных питательных средах, например, на кровяном агаре. Их роль в патологии человека и животных установлена в конце ХIХ столетия, когда первые бактероиды были изолированы из очагов некротических поражений. Вместе с тем они являются едва ли не главным компонентом нормальной микрофлоры, количественно доминируя в ротовой полости, толстом кишечнике и влагалище.
Семейство Bacteroidaceae включает более десятка родов, из которых в медицинской микробиологии наиболее известны представители Bacteroides, Fusobacterium и Leptotrichia. Недавно утверждены новые родовые таксоны: Porphyromonas, Previtella, Selemonas, Wolinella. Род Bacteroides включает наибольшее число видов и играет основную роль в формировании нормальной микрофлоры и в патологии человека. Именно поэтому за всем семейством утвердилось название Bacteroidaceae, хотя в более узком смысле бактероидами следует называть представителей единственного рода — Bacteroides.
Классификация
Базисная дифференцировка бактероидов (речь идет о всем семействе Bacteroidaceae) основана на различиях короткоцепочечных жирных кислот (уксусная, сукциновая, n-бутировая, молочная), образующихся из белков и углеводов, на способности к пигментообразованию и структурных особенностях пептидогликана. Например, фузобактерии отличаются характерной морфологией (длинные веретенообразные палочки с заостренными концами), хотя многое зависит от условий культивирования (рис. 1).


Несмотря на дополнения и поправки, внесенные за последнее время в классификационные схемы, таксономику бактероидов нельзя признать завершенной.
Роды Bacteroides, Porphyromonas, Previtella. Это весьма неоднородная группа, чему соответствует широкий разброс в содержании гомологичных пар пурин-пиримидиновых оснований ДНК разных видов: показатель «гуанин+цитозин» колеблется в пределах 28—61%. Не случайно за родом Bacteroides закреплено более 40 видов, которые принято делить на 3 основные группы: fragilis, melaninogenicus-oralis и asaccharоlyticus (табл. 1). Род Bacteroides предложено расчленить на три самостоятельных родовых таксона: Bacteroides (собственно бактероиды), Porphyromonas и Previtella (см. ниже).
Таблица 1
Виды рода Bacteroides
| Группа fragilis | Группа melaninogenicus-oralis | Группа asaccharolyticus |
| B. fragilis*** B. vulgatus B. distasonis B. merdae B. caccae B. ovatus B. thetaiotamicron* B. eggerthii B. uniformis B. stercoris B. splanchnicua |
B. melaninogenicus B. loescheii B. denticola B. intermedius** B. corporis B. levii B. oris* B. buccae* B. zoogleoformans B. buccalis B. veroralis B. heparinolyticus B. macacae B. oralis B. oulorum B. bivius* B. disiens* |
B. asaccharоlyticus** B. gingivalis** B. endodontalis* B. urealyticus* B. gracilis* B. putredinis B. praeacutus B. pneumosintes B. coagulans B. nodosus |
* степень патогенности для человека.
Группа fragilis. Типовой вид B. fragilis открыт в конце XIX в. В состав группы входят бактероиды, обладающие рядом одинаковых признаков и общей экологией: все они являются комменсалами толстого кишечника, основным элементом его нормальной микрофлоры. Представляют собой мелкие, неподвижные палочки и коккобациллы с умеренным плеоморфизмом. Отличительными признаками служат устойчивость к пенициллину и стимуляция роста 20% желчью. Несмотря на фенотипическое и экологическое сходство, отдельные виды связаны слабым генетическим родством (низкий процент гомологии ДНК). Некоторые из них (прежде всего B. fragilis) вызывают деструктивные процессы, главным образом в брюшной полости, куда они попадают при повреждении кишечника.
Группа melaninogenicus-oralis. Название группы предложено в 1921 г. после сообщения о грамотрицательных анаэробных палочках, образующих на кровяном агаре колонии черного цвета. Позже было установлено, что пигментация связана с производными гемоглобина, и все бактероиды, наделенные этим признаком, стали относить к группе melaninogenicus. В дальнейшем оказалось, что образование пигментированных колоний нельзя считать надежным классификационным критерием, и в эту группу были включены виды, близкие к B. melaninogenicus по метаболическому профилю (продукция n-бутировой кислоты в качестве основного конечного метаболита), но лишенные способности к пигментообразованию. Эпитет «oralis» означает, что главным биотопом для бактероидов данной группы является ротовая полость; только два вида (B. bivius и B. disiens) входят в состав нормальной микрофлоры влагалища.
Большинство бактероидов этой группы представляют плеоморфные короткие палочки или коккобациллы. Виды дифференцируют по пигментообразованию и стандартным биохимическим тестам (продукция индола, гидролиз эскулина, ферментация углеводов). По современной классификации 13 видов этой группы выделены в род Previtella (табл. 2).
Таблица 2
Виды родов Porphyromonas и Previtella
| Новое название | Прежнее название |
|
Porphyromonas asaccharоlytica P. endodontalis P. gingivalis Previtella bivia P. buccae P. buccalis P. denticola P. disiens P. heparynolitica P. intermedia P. melаninogenica P. oralis P. oris P. oulorum P. ruminicula P. veroralis |
Bacteroides asaccharо-lyticus B. endodontalis B. gingivalis Bacteroides bivius B. buccae B. buccalis B. denticola B. disiens B. heparynolyticuis B. intermedius B. melаninogenicus B. oralis B. oris B. oulorum B. ruminicula B. veroralis |
Группа asaccharolyticus. Сюда относятся бактероиды, не образующие кислоту из глюкозы и других углеводов (они способны утилизировать их без брожения). В целом это неоднородная (в том числе экологически) категория с неясными таксономическими контактами. Некоторые из них образуют пигмент на кровяном агаре и ранее относились к группе melaninogenicus. Входят в состав нормальной микрофлоры ротовой полости (большинство видов), толстого кишечника и, возможно, влагалища (B. asaccharolyticus). Три вида этой группы выделены в род Porphyromonas (см. табл. 2).
Род Fusobacterium. Неодинаковые по размеру и форме бактерии. В патологическом материале выглядят как кокки, палочки и длинные (до 80—100 мкм) нити. Для некоторых видов характерно образование L-форм и сферопластов, из-за чего ранее их классифицировали как род Spherophorus. В чистых культурах картина гораздо гомогеннее: прямые или искривленные палочки и короткие нити с заостренными концами, напоминающие веретено (см. рис. 1). Отсюда и название — фузиформные бактерии (лат. fusiformis — веретенообразный), которое появилось в конце ХIХ в. Род включает более 10 видов, изолированных из нормальной ротовой полости человека и животных, а также патологического материала (очагов некроза) (табл. 3).

Еще раньше лептотрихии называли бактериями (фузобактериями) Винсента, так как с момента открытия (Vincent, 1896) их считали возбудителем ангины Винсента (фузоспирохетоза) — заболевания, напоминающего дифтерию зева. В настоящее время лептотрихии рассматриваются как элемент нормальной микрофлоры ротовой полости, а их патогенность ставится под сомнение. Приоритет в этиологии ангины Винсента отдается истинным фузобактериям (F. nucleatum), превителлам (Р. intermediа) и спирохетам (Borrelia vincentii).
Представительство в нормальной микрофлоре
Бактероиды (мы используем этот термин как собирательное понятие для всех грамотрицательных бесспоровых анаэробных палочек) принадлежат к ведущим компонентам нормальной микрофлоры слизистых оболочек полости рта, толстого кишечника и влагалища, где их количество во много раз превышает содержание аэробных и факультативно-анаэробных бактерий. Виды, населяющие эти участки тела, различны и формируют дискретные популяции.
Ротовая полость. Главным биотопом являются десневые карманы и субдесневые бляшки. У детей до смены молочных зубов бактероиды обнаруживаются непостоянно, но затем их количество возрастает, а присутствие становится обязательным. Кроме собственно бактероидов здесь обнаруживаются представители родов Previtella, Porphyromonas, Selenomonas, Wolinella, фузобактерии и лептотрихии. Бактероиды fragilis-группы в ротовой полости отсутствуют.
Количество фузиформных бактерий, регистрируемое в десневой микрофлоре путем прямой микроскопии, в тысячи раз превосходит число культивируемых клеток. Многие из изолятов по своим свойствам не укладываются в современные классификационные схемы и условно объединяются в довольно расплывчатый вид Fusobacterium nucleatum (polymorphum).
При гингивитах содержание бактероидов в ротовой полости увеличивается.
Кишечник. Практически вся микрофлора кишечного тракта здоровых людей представлена бактериями толстого кишечника. Подсчитано, что их более 400 видов, причем около 99% составляют анаэробы. На кишечную палочку приходится не более 0,01% всей бактериальной массы; других аэробных и факультативно-анаэробных бактерий еще меньше. Большинство анаэробов составляют бактероиды и бифидобактерии (примерно в равных пропорциях). Около 80% грамотрицательных анаэробов относятся к fragilis-группе, хотя (и это принципиально!) ее патогенетический лидер, B. fragilis, присутствует в небольшом количестве. Доминируют B. vulgatus, B. thetaiotaomicron, B. distasonis и B. eggerthii. Достаточно много асахаролитических бактероидов (преобладает Porphyromonas asaccharоlytica). Представители группы melaninogenicus-oralis (включая Previtella) и фузобактерии встречаются во много раз реже.
Бактероиды играют важную роль в физиологии толстого кишечника. Они, например, метаболизируют желчные кислоты, готовя их к кишечно-печеночному циклу. Это может иметь и негативные последствия, так как в ходе микробзависимой трансформации желчных кислот образуются канцерогенные метаболиты (циклопентанфенантрены). Замечено, что вероятность рака толстой кишки связана с типом питания и процентным содержанием грамотрицательных анаэробов в фекалиях.
Влагалище. У большинства женщин в цервикальном отделе влагалища присутствуют бактероиды группы melaninogenicus-oralis (Previtella). Преобладающим видом является P. bivius; достаточно типичны P. disiens, P. intermedius и P. melaninogenicus. Из бактероидов асахаролитической группы встречается Porphyromonas asaccharоlytica. Бактероиды fragilis-группы и фузобактерии выделяются редко. Повышение количества бактероидов fragilis-группы, особенно B. fragilis, говорит о патологии генитального тракта.
Патогенность: факторы и реализация
Представления о болезнетворности бактероидов возникли в конце ХIХ в. после выделения от людей и животных с деструктивными гнилостными процессами. Картина уточнялась по мере совершенствования техники изучения облигатных анаэробов и утверждения принципов их классификации. Сегодня бактероидная патология признается одной из важных проблем клинической микробиологии. Чаще это эндогенная инфекция, связанная с повреждением слизистых оболочек, где в изобилии вегетируют различные бактероиды. Они атакуют глубокие ткани, вызывая более или менее обширные гангренозные поражения, которым сопутствует гнилостный запах, а иногда газообразование. Развитию процесса благоприятствует гипоксия из-за недостаточного кровоснабжения. Поэтому бактероиды особенно активно размножаются в поврежденных и некротизированных тканях, а также в зонах ишемии, например при нарушении периферийного кровообращения. Тканевая деструкция, нарастающая по мере инфекции, усиливает недостаток кислорода, стимулируя размножение анаэробов. Этому способствуют инородные тела, а также то, что в очагах поражения бактероиды обычно соседствуют с аэробными и факультативно-анаэробными бактериями, которые, поглощая кислород, усиливают анаэробиоз. Чувствительность к бактероидам повышается при смешанном инфицировании животных, например, при совместном заражении бактероидами и кишечной палочкой. Смешанный характер клинических инфекций объясняется тем, что при повреждении слизистых оболочек в ткани попадают все бактерии, населяющие данный участок тела. При углублении гипоксии анаэробы становятся ведущим фактором деструктивного (некротического) процесса.
К счастью, лишь немногие бактероиды обладают патогенностью. Из них только один вид, B. fragilis, способен вызывать моноинфекции; остальные встречаются в ассоциациях между собой и с другими бактериями. Болезнетворность бактероидов и фузобактерий (лептотрихии, по-видимому, безвредны) напрямую не связана с их количественным содержанием в нормальной микрофлоре, т.е. с вероятностью массивного обсеменения тканей. Более того, их лидер, B. fragilis, значительно уступает по численности более безобидным видам. Это означает, что патогенные (точнее условно-патогенные) бактероиды наделены особыми свойствами, которые обеспечивают их агрессивность. Учитывая смешанный характер большинства инфекций, когда в очаге поражения одновременно присутствуют различные бактероиды и/или фузобактерии, можно говорить об их интегративном патогенетическом потенциале, который складывается из суммы факторов нескольких видов бактерий. При моноинфекции агрессивности часто не хватает, и процесс не получает развития.
Как уже говорилось, бактероидная инфекция обычно начинается с повреждения слизистых оболочек, когда нормальная микрофлора получает доступ в глубжележащие ткани. Яркий пример — перфорация кишечника, ведущая к перитониту. Это результат совместной аэробной (факультативно-анаэробной) и анаэробной инфекции, где бактероидам (прежде всего B. fragilis) принадлежит главная роль. Первым негативным для себя фактором, с которым анаэробы сталкиваются в новой среде обитания, яв-ляется кислород. Большинство бактероидов достаточно терпимы к кислороду и по крайней мере несколько часов выживают в обычной атмосфере. Мы уже говорили о ситуациях, которые ведут к местной гипоксии, способствуя стабилизации анаэробного инфекта. Но ускользание от кислорода может быть и активным. Некоторые бактероиды располагают ферментами, которые разрушают высокотоксичные метаболиты кислорода. Такой способностью обладает, в частности, B. fragilis, продуцирующий каталазу и супероксиддисмутазу. Поэтому уже на первом этапе (сразу после пассивной инвазии в ткани) происходит селекция бактерий по устойчивости к кислороду. Вероятность продолжения процесса наиболее реальна для аэротолерантных штаммов.
Для отражения фагоцитарной атаки некоторые из бактероидов располагают полисахаридной капсулой, которая защищает их от фагоцитоза, способствуя абсцедированию и системной инвазии. Этот признак опять-таки лучше всего выражен у B. fragilis, подчеркивая патогенетическое лидерство данного вида среди бактероидов. Впрочем, и авирулентные штаммы могут обретать способность к капсулообразованию и индуцировать абсцессы при смешанной инфекции с другими аэробными и анаэробными бактериями.
Еще один антифагоцитарный фактор — низкомолекулярные жирные кислоты. Они продуцируются бактероидами и фузобактериями и отрицательно влияют на функции многих клеток, в том числе фагоцитов. Так, бактероиды fragilis-группы накапливают значительное количество сукциновой (янтарной) кислоты, которая подавляет кислородзависимую бактерицидность и хемотаксис нейтрофилов.
Наконец, бактероиды располагают протеолитическими ферментами, разрушающими иммуноглобулины и факторы комплемента. Это помогает им и соседствующим с ними бактериям противостоять опсонофагоцитозу, который и без того ослаблен из-за низкого притока крови в зоны ишемии.
Прямой некротизирующий эффект, столь типичный для бактероидов и фузобактерий, связан с действием их гистолитических ферментов. Они расщепляют тканевые структуры, способствуя бактериальной инвазии и превращая ее в расползающуюся или абсцедирующую зону гангренозного распада. К таким ферментам относятся разнообразные протеиназы (коллагеназа, фибринолизин и др.), липазы, нейраминидаза, гиалуронидаза, нуклеазы, хондроитинсульфатаза и пр. Некоторые из бактероидов продуцируют гепариназу — фермент, который, расщепляя гепарин, способствует тромбообразованию, усиливая тканевую ишемию и, следовательно, анаэробиоз.
Подобно всем грамотрицательным бактериям, бактероиды содержат липополисахаридный эндотоксин. Его биологическая активность неодинакова у разных видов, а в ряде случаев близка к нулю. Инертным липополисахаридом располагает, в частности, B. fragilis, что, однако, не мешает ему лидировать по патогенности. Дело в том, что основные эффекты эндотоксина проявляются после проникновения в кровь и опосредованы через систему эндогенных медиаторов воспаления. Он не токсичен в зоне локального поражения и в этом отношении принципиально отличается от агентов с прямым повреждающим эффектом, которые доминируют при анаэробной инфекции.
В табл. 4 суммированы факторы, с которыми связана болезнетворность бактероидов. Повторим, что они неодинаково выражены у разных видов и даже штаммов, но благодаря смешанному характеру инфекций могут присутствовать в очаге поражения. Эталоном является B. fragilis, на изучении которого базируются основные представления о механизмах патогенности бактероидов.
Таблица 4
Факторы патогенности бактероидов
| Факторы | Патогенетический эффект |
| Гистолитические ферменты (протеиназы, галуронидаза, липазы, нуклеазы и пр.) | Деструкция тканей, матриксных белков (коллагена, фибронектина, ламинина), фибрина Подавление иммунитета (деструкция иммуноглобулинов, комплемента) |
| Ферменты, инактивирующие кислород (каталаза, супероксиддисмутаза) | Выживание в аэробных условиях (аэротолерантность) |
| Капсульный полисахарид | Подавление фагоцитоза Абсцедирование |
| Лектиноподобные поверхностные белки, фимбрии (?) | Адгезия (мезотелиальные, эпителиальные клетки) |
| Липополисахаридный эндотоксин | Синдром эндотоксинемии |
| Гепариназа | Локальные тромбозы и тканевая ишемия |
| Жирные кислоты | Антифагоцитарный эффект |
| IgA-протеаза | Подавление иммунитета слизистых оболочек (разрушение секреторных антител) |
| Бета-лактамазы | Устойчивость к антибиотикам |
Патология
Большинство инфекций начинается в тканях, прилегающих к зонам естественного обитания бактероидов, т.е. к слизистым оболочкам ротовой полости, кишечника и полового тракта (у женщин). Отсюда следует, что видовой спектр возбудителей в очагах поражения должен быть неодинаковым, отражая особенности микрофлоры соответствующих участков тела. Так, абдоминальные и перинеальные инфекции обычно связаны с fragilis-группой и асахаролитическими разновидностями (Porphyromonas asaccharоlytica), причем в 60—70% случаев выделяется B. fragilis. В патологии женских гениталий наряду с B. fragilis существенную роль играют B. bivius, P. disiens, P. melaninogenica. В оральной патологии преобладают бактероиды родов Previtella, Porphyromonas и фузобактерии. Впрочем, к бактероидной и фузобактериальной инфекциям чувствительны любые ткани, и благодаря контактному, лимфогенному и гематогенному метастазированию очаги деструктивного поражения могут возникать в различных органах.
Инфекции ротовой полости и зубов. Большинство инфекций возникает из десневой микрофлоры. В норме количество бактероидов и фузобактерий здесь невелико, но значительно возрастает при гингивитах. Бактероиды активно включаются в поражение пульпы кариозных зубов (они входят в состав микрофлоры зубных бляшек), корневых каналов, периодонта. Такие инфекции могут осложняться прикорневыми гранулемами, одонтогенными абсцессами, флегмоной лица и шеи (ангина Людвига, нома), челюстным остеомиелитом. Классической патологией, ассоциированной с бактероидами, является фузоспирохетоз — некротический язвенный гингивит, иногда в сочетании с гангренозным фарингитом (ангина Винсента). В его основе лежит смешанная инфекция, связанная с фузобактериями (F. nucleatum), превителлами (Р. intermediа) и спирохетами (Воrrelia vincentii) (рис. 2).

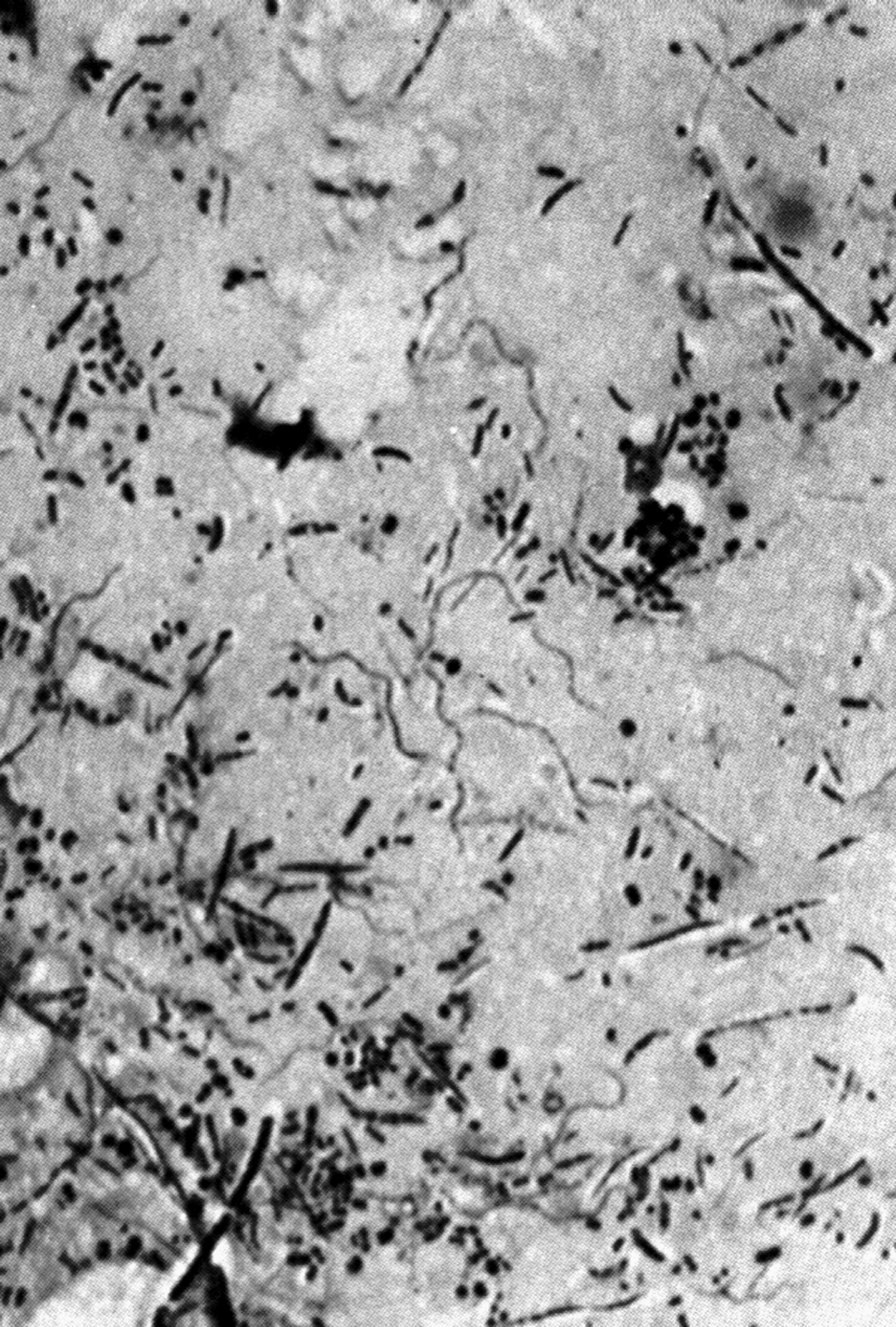
Ведущая роль в развитии периодонтита принадлежит бактериодам родов Porphyromonas (P. gingivalis), Previtella (P. intermedia), Fusоbacterium (F. nucleatum), Wolinella (W. recta). Наибольший интерес вызывает P. gingivalis. Бактероиды этого вида располагают протеолитическими ферментами, которые готовят участки для колонизации других бактерий, обнажая адгезивные сайты на молекулах коллагена и фибронектина. Кроме того, они активно внедряются в эпителиальные клетки и, укрываясь от действия антибактериальных факторов, запускают повреждение изнутри.
Абдоминальные инфекции. Бактероиды участвуют в формировании смешанных очагов инфекции при интраабдоминальном сепсисе, который развивается после прободения кишечника. Если на этапе раннего перитонита (очагового или разлитого) доминируют колиформные бактерии (прежде всего кишечная палочка), то после абсцедирования лидерство переходит к бактероидам (чаще B. fragilis), хотя инфекция всегда остается смешанной. Двухфазность интраабдоминального сепсиса находит отражение в клинической картине, требуя грамотного подбора антибиотиков для суммарного воздействия на оба компонента инфекции — колиформные бактерии и бактероиды.
С бактероидами связана еще одна проблема. Среди B. fragilis обнаружены энтеротоксигенные штаммы, вызывающие диарею у человека, особенно у детей. Действующим началом является токсин (фрагилизин), который относится к цинксодержащим металлопротеиназам (мецинцинам). Он нарушает структурно-функциональные контакты (парацеллюлярный барьер) между эпителиоцитами кишечника, вызывая их эксфолиацию и гиперсекрецию жидкости. Повреждение парацеллюлярного барьера открывает доступ бактериям в глубжележащие ткани, запуская воспалительную реакцию.
Инфекции женского полового тракта. Являясь частью нормальной микрофлоры влагалища, бактероиды могут активироваться на фоне сопутствующей патологии, например при злокачественных опухолях (после местной лучевой терапии) и хирургических травмах. Они включаются в развитие пиогенных инфекций матки и таза, таких как эндометрит, параметрит, сальпингит, тубоовариальные абсцессы, пельвиоперитонит, постабортальные, послеродовые и постхирургические флегмоны.
Бактероиды группы melaninogenicus-oralis (чаще Previtella bivia) имеют отношение к так называемому вагинозу — патологии влагалища, которая протекает без воспаления (отсюда вагиноз, а не вагинит), сопровождается повышением рН вагинальной среды (более 4,5) и дурно пахнущими (но не гнойными) выделениями (рыбный запах аминов, продуцируемых бактероидами). Поражение связано по меньшей мере с двумя группами бактерий — бактероидами и гарднереллами (Gardnerella vaginalis). G. vaginalis выделяется от 20—40% здоровых женщин, но присутствует в уретре большинства мужчин и может передаваться половым путем. Возможно, что именно заражение мужскими гарднереллами нарушает баланс в вагинальном микробиоценозе и создает условия для избыточного размножения анаэробов. Об участии последних говорит то, что болезнь хорошо лечится метронидазолом, к которому чувствительны бактероиды. При микроскопии в выделениях находят эпителиальные клетки, усыпанные грамотрицательными коккобациллами (англ. clue cells, т.е. клетки, дающие ключ к разгадке). Этот признак используется в дифференциальной диагностике вагинальных инфекций (рис. 3).

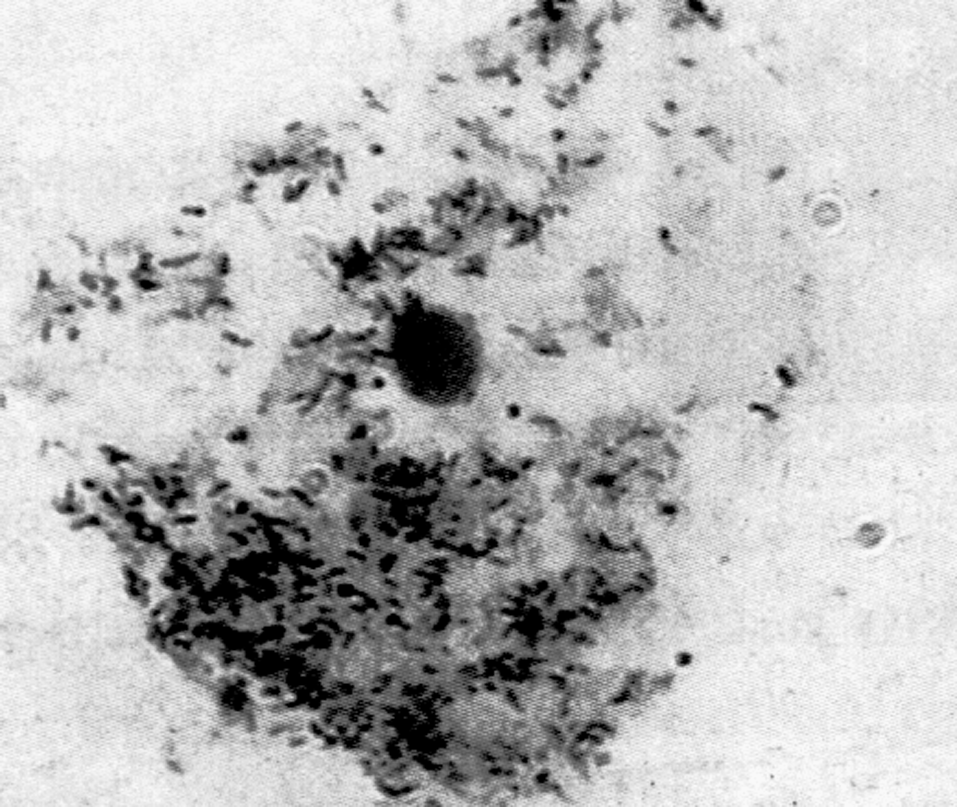
Инфекции ЛОР-органов и респираторного тракта. Кроме отмеченной выше ангины Винсента, оральные бактероиды и фузобактерии могут быть причиной перитонзиллярных и ретрофарингеальных абсцессов, а также перитонзиллярной флегмоны (целлюлита). Их присоединение к хроническому тонзиллиту проявляется симптомом гнилостного дыхания.
В доантибиотическую эру F. necrophorum был достаточно обычной причиной острого среднего отита, мастоидита и синуситов, которые иногда осложнялись тяжелой бактериемией. Сегодня анаэробы редко встречаются при острой ЛОР-патологии, но для хронических процессов они весьма типичны. Виды, ассоциированные с хроническим средним отитом и синуситами, происходят из оральной микрофлоры, хотя нередко (примерно в половине случаев) выделяется и B.fragilis. У половины больных хроническим гнойным отитом содержание анаэробов в очаге поражения (обычно в сочетании с другими бактериями) достигает 109/мл.
В составе смешанных инфекций представители бактероидов служат одной из главных причин деструктивной (некротической) пневмонии, абсцессов легких, эмпиемы плевры, прогрессирующих бронхоэктазов. Они проникают в легкие аспирационным путем из ротовой полости и верхних дыхательных путей, чаще как осложнение периодонтита, перитонзиллярных инфекций, хронического синусита, среднего отита и пр. Это объясняет спектр бактероидов, выделяемых при легочно-плевральной патологии: доминируют оральные бактероиды (группа melaninogenicus-oralis, Previtella) и фузобактерии (обычно F. nucleatum); B. fragilis встречается редко (чаще при эмпиеме плевры).
Инфекции центральной нервной системы. Бактероиды относятся к наиболее типичным возбудителям абсцессов головного мозга, особенно таких, которые возникают при распространении инфекции из соседних тканей (хронический средний отит, мастоидит, синуситы). В смешанной микрофлоре очагов подобного типа бактероиды и фузобактерии встречаются с наибольшим постоянством и доминируют в количественном отношении. Лидирует B. fragilis, но нередко присутствуют оральные бактероиды и F. nucleatum. Возможны и метастатические абсцессы, возникающие при заносе инфицированных тромбов из отдаленных очагов поражения. Такие абсцессы обычно развиваются как моноинфекция, бактериальный профиль которой отражает микрофлору источника метастазов. Так, причиной метастатических абсцессов ЦНС при плевропульмональных инфекциях нередко служат фузобактерии; B. fragilis выделяются редко. Одонтогенные абсцессы чаще связаны с превителлами, порфиромонами и фузобактериями.
Инфекции кожи и мягких тканей. Попав в подкожную клетчатку, бактероиды включаются в развитие флегмонозного процесса (в зарубежной литературе чаще пользуются термином целлюлит), который может захватить обширную территорию, а при закупорке сосудов перейти в гангрену. Инфекция, как правило, смешанная, причем именно представители бактероидов играют главную роль в повреждении тканей. Характерен гнилостный запах, а тромбоз подкожных артериол и венул ведет к ишемии и некрозу, способствуя размножению анаэробов и быстрому развитию процесса.
Тяжелейшей разновидностью анаэробной флегмоны является нома (от греч. nomes — разъедание, распространение) — быстро прогрессирующая гангрена лица, которая начинается в зоне переходной складки кожи и слизистой губ. Ее принято считать фузоспирохетозом, хотя не исключено, что главную роль играют оральные бактероиды. Болезнь чаще наблюдается у детей, ослабленных тяжелой инфекцией (корь, скарлатина и пр.) либо истощенных из-за недостатка питания.
Некротизирующая флегмона, которая поражает наружные половые органы, быстро распространяясь на соседние области, называется гангреной Фурнье. Она начинается как раневая инфекция (после операций в паховой области, промежности или нижнем отделе живота) или как следствие незначительных повреждений гениталий. Инфекция смешанная, но лидерство принадлежит бактероидам.
Язвы при варикозном расширении вен, диабете и пролежнях (decubitus ulcer) почти всегда инфицированы бактероидами и фузобактериями. Они обнаруживаются в глубине поражений; поверхность колонизирована аэробными и факультативно-анаэробными бактериями.
Есть данные, что бактероиды выделяются от 70% больных паронихией и вызывают осложнения после укусов животных (оральные бактероиды и фузобактерии).
Бактериемия. Как уже говорилось, бактероиды и фузобактерии могут попадать в кровь, создавая опасность гематогенных метастазов. Известны случаи эндокардита, связанные с анаэробными бактериями, в том числе с бактероидами, но это наблюдается крайне редко. По системным эффектам (септический синдром) бактероидная бактериемия значительно уступает энтеробактериальной. Это, по-видимому, объясняется низкой биологической активностью эндотоксина главных в патогенетическом отношении видов бактероидов.
Принципы диагностики и этиотропной терапии
Выделение, идентификация и сохранение чистых культур бактероидов представляют большие трудности и являются дорогостоящими процедурами. Это объясняется необходимостью поддерживать анаэробные условия на всех этапах анализа, начиная с получения клинического материала и далее при транспортировке, культивировании и т.д. По подсчетам зарубежных коллег, видовая идентификация бактероидов (например, B. fragilis) обходится в сто раз дороже, чем анализ на энтеробактерии, например на ту же кишечную палочку. Требуется специальное оборудование, оснащенное устройствами для поддержания бескислородной атмосферы, а это под силу немногим лабораториям. К тому же патологический материал часто загрязнен случайной микрофлорой, не имеющей отношения к заболеванию. В связи с этим предложено использовать выявление бактероидов на основе определения микробных метаболитов — летучих жирных кислот (уксусная, пропионовая, масляная, капроновая и др.), а также ароматических производных (фенилпропионовая и фенилуксусная кислоты, метаболиты фенола, индола, пиррола и др). С этой целью применяют газово-жидкостную хроматографию, результаты которой хорошо коррелируют с бактериальной обсемененностью пораженных тканей и опасностью местных и общих осложнений. Одним из решений проблемы может быть выявление генетических маркеров инфекции на основе полимеразной цепной реакции (см. «Диагностика инфекционных заболеваний»).
За редкими исключениями бактероиды чувствительны к метронидазолу в концентрациях, легко достижимых внутри организма при введении обычных дозировок. Первоначально метронидазол был предложен как антипротозойный антибиотик (например, против Giardia lamblia, Entamoeba histolytica и Trichominas vaginalis). Его биоцидность для анаэробных грамотрицательных бактерий и клостридий была обнаружена позднее. Он не действует на аэробные и факультативно-анаэробные бактерии. Это связано с тем, что антибактериальная активность метронидазола (он ингибирует синтез ДНК) проявляется лишь после частичного восстановления его нитрогрупп. Это умеют делать анаэробы и лишь изредка аэробные бактерии.
Большинство бактероидов чувствительны к линкозамидам, хотя есть сообщения о появлении устойчивых штаммов. Клиндамицин (в сочетании с метронидазолом) до сих пор широко применяется для лечения анаэробных инфекций.
Все представители бактероидов устойчивы к аминогликозидам. Поэтому их используют для приготовления элективных сред с целью подавления посторонней микрофлоры.
С бета-лактамными антибиотиками дело обстоит сложнее. Бактероиды fragilis-группы не чувствительны к пенициллину. Это связано с конститутивной бета-лактамазой, обладающей широким спектром активности против пенициллинов и цефалоспоринов. Большинство представителей группы melaninogenicus-oralis (в том числе превителлы) поначалу были чувствительны к пенициллину, который считался антибиотиком выбора для лечения смешанных аэробно-анаэробных инфекций, локализованных выше диафрагмы. Но сегодня благодаря плазмидозависимой бета-лактамазе (она отличается от бета-лактамазы fragilis-группы) около половины клинических изолятов группы melaninogenicus-oralis устойчивы к пенициллину. Кроме того, в очагах смешанной инфекции источником бета-лактамаз могут быть аэробные бактерии, обеспечивающие таким путем косвенную защиту анаэробов от антибиотиков.
В отличие от бактероидов, фузобактерии и лептотрихии чувствительны к терапевтическим дозировкам пенициллина. Некоторые из новых цефалоспоринов обладают активностью против бактероидов, но их эффект ниже, чем у клиндамицина и метронидазола. Отмечается высокая чувствительность к имипенему и хлорамфениколу. К азтреонаму и монобактаму бактероиды резистентны. Тетрациклин и его аналоги, которые некогда были высокоактивны против бактероидов, потеряли свою эффективность, и в настоящее время к ним устойчивы более половины штаммов.
В целом сегодня имеется несколько препаратов, достаточно эффективных против анаэробной инфекции. Обычно используют комплекс антибиотиков, учитывая смешанный характер инфекции и необходимость одновременного воздействия на весь спектр аэробных и анаэробных бактерий в очаге поражения. Кроме того, как и при любой анаэробной инфекции, большое (нередко решающее) значение имеет хирургическая обработка ран с иссечением всех нежизнеспособных тканей.
Эшерихии
Нет стада без бодливой коровы.
- Экологическая стратегия.
- Иммунотипирование.
- Серовары и патология.
- Многоликость диареегенных эшерихий.
- Внекишечные септические инфекции.
Род Escherichia включает несколько видов, но безоговорочным лидером среди них является кишечная палочка — E. coli. Это типовой вид семейства Entеrobacteriaceae, объединяющего грамотрицательные факультативно-анаэробные палочковидные бактерии, которые не образуют спор и обладают рядом других общих признаков.
В родовом названии закреплен приоритет немецкого бактериолога T. Эшериха, выделившего кишечную палочку в 1885 г. из фекалий ребенка с «детской холерой», которая была широко распространена в Европе. Но признание ее болезнетворности задержалось на долгие годы. Дело в том, что сходные бактерии неизменно выявлялись в кишечнике здоровых детей и взрослых, что противоречило одному из постулатов Коха, жестко различавших патогенные и безвредные микробы. Убиквитарность кишечной палочки зафиксирована и в первом названии, предложенном Т. Эшерихом — Bacterium coli communis. Ее роль в патологии кишечника не отрицалась, но у многих вызывала сомнения. Более убедительным выглядело участие кишечной палочки во внекишечной патологии, особенно при воспалительных заболеваниях мочевыводящей системы. Лишь много позже при помощи антигенных маркеров, недоступных современникам Т. Эшериха, удалось расшифровать парадокс кишечной палочки, доказав патогенетические потенции ее отдельных представителей.
Современный интерес к E. coli выходит далеко за рамки медицинской микробиологии. С кишечной палочкой связаны едва ли не главные достижения молекулярной биологии и генетики. Вместе со своими плазмидами и бактериофагами она служит «рабочей лошадкой» генной инженерии и широко используется для получения искусственных (рекомбинантных) белков.
Видовая идентификация эшерихий не представляет трудностей. Подобно другим энтеробактериям, они неприхотливы и культивируются на простейших питательных средах, содержащих глюкозу (источник углерода) и неорганический азот. В отличие от многих энтеробактерий кишечная палочка быстро сбраживает лактозу. Это легко улавливается на средах, которые содержат лактозу и индикатор, меняющий цвет при закислении питательной среды. Колонии принимают окраску соответствующего индикатора и легко отличимы от колоний бактерий, не ферментирующих или слабо ферментирующих лактозу. Следует лишь помнить, что кроме стандартных встречаются атипичные штаммы, не активные или малоактивные в отношении лактозы. Их приходится дифференцировать в группе так называемых колиформных бактерий, к которым некогда относили все энтеробактерии за исключением шигелл, сальмонелл, протея и йерсиний. Современное фенотипирование, подкрепленное генетическими маркерами, позволяет при желании отойти от утилитаризма, доведя идентификацию хотя бы до родового уровня заметно разросшегося семейства энтеробактерий (согласно последней классификации, оно включает 29 родовых таксонов). Вместе с тем понятие колиформности сохраняет практический смысл и сегодня, особенно в тех случаях, когда не требуется тонкой идентификации штаммов. Кроме кишечной палочки, к колиформным бактериям относят представителей родов Klebsiella, Enterobacter, Citrobacter, Arizona, Serratia, Providencia. Они вносят определенный (хотя и гораздо меньший) вклад в инфекционную патологию прежде всего как возбудители септических госпитальных инфекций.
Экология кишечной палочки типична для энтеробактерий: ее резервуаром служит дистальный отдел кишечника человека и теплокровных животных (рис. 1).

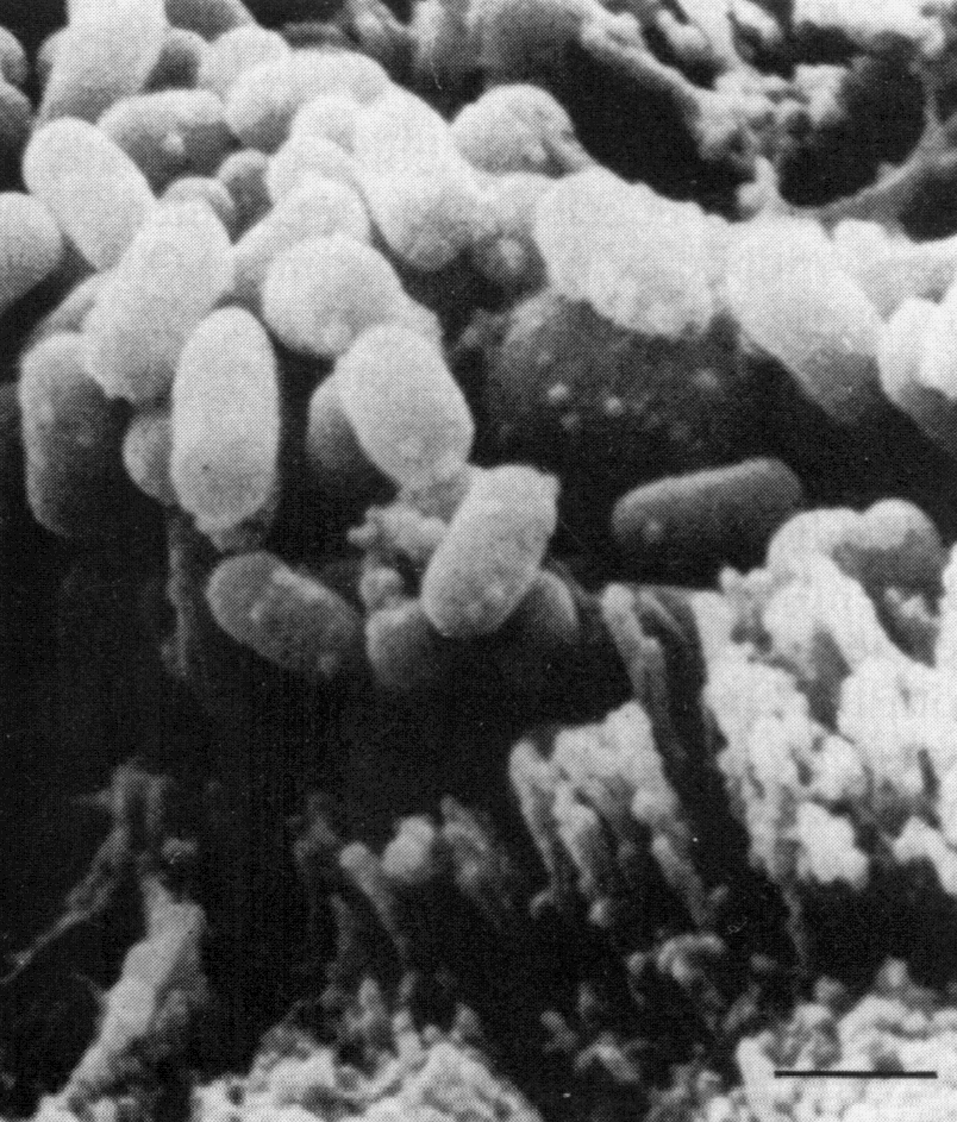
Кроме того, эшерихии обнаруживаются в кишечном содержимом пресмыкающихся, рыб и насекомых. E. coli является символом нормальной микрофлоры и до утверждения представлений о количественном доминировании облигатных анаэробов прочно удерживала здесь лидирующие позиции. Эшерихии растут в режиме от 10 до 46°С (оптимум 37°С), ограничивая питательные потребности неорганическим азотом и примитивными источниками углерода (углеводы, ацетат натрия и пр.). Это обеспечивает широкие возможности для выживания во внешней среде, хотя сапрофитизм эшерихий факультативен и для продолжения жизненного цикла им необходим кишечник человека или животных.
Широкое представительство в нормальной микрофлоре служило поводом для признания безвредности кишечной палочки. Попытки изменить положение, признав за эшерихиями патогенетическую неоднородность, воспринимались с интересом, но не вели к продуктивным обобщениям. Радикальный сдвиг произошел в 1950-х гг. после того, как датский бактериолог Ф. Кауфман обосновал принципы серологической классификации (иммунотипирования) эшерихий на основе поверхностных антигенов. Серологическими маркерами кишечной палочки (как и других энтеробактерий) являются О-, К- и Н-антигены. Их носителями служат соответственно липополисахарид наружной мембраны, микрокапсула и жгутики. По данным иммунотипирования различных штаммов (его проводят в реакции агглютинации) дифференцируют более 170 вариантов О-антигена, 90 — К-антигена и 50 — Н-антигена.
Главным является деление на О-типы (группы), принадлежность к которым загадочным образом связана с вирулентностью. К-антигены по физико-химическим свойствам (устойчивость к прогреванию, этанолу, формалину и пр.) подразделяются на три категории — L, A и В. В большинстве это кислые полисахариды, экранирующие О-антиген в реакции агглютинации: многие К-позитивные штаммы не агглютинируются О-антисыворотками. Это вносило путаницу, которую удалось преодолеть путем избирательного разрушения К-антигенов (кипячение). Каждый штамм располагает единственным К-антигеном, нередко ассоциированным с определенной О-группой. Некоторые из К-позитивных эшерихий обладают повышенной вирулентностью, так как более устойчивы к фагоцитозу и бактерицидному действию комплемента.
Полное иммунотипирование завершается определением Н-антигена. Это белок, который имеется только у подвижных (жгутиковых) штаммов и может быть слабо выражен в первичных культурах. Корреляция между Н- и О-типами выражена слабо.
Каждый иммунофенотип (на практике пользуются понятиями «серотип», или «серо-вар») имеет О:К:Н-формулу (например, О8:К25:Н9), причем благодаря случайности в комбинациях О-, К- и Н-антигенов число возможных серотипов превышает 2000. Обычно выявляют лишь О-групповую принадлежность, так как именно этот признак лучше всего согласуется с патогенностью (хотя и не объясняет ее!).
Штаммовое многообразие в сочетании с реактивностью хозяина определяют три главных варианта эколого-патогенетической стратегии кишечной палочки:
- непатогенные (резидентные) комменсалы, пожизненно колонизирующие толстый кишечник;
- эшерихии, вызывающие внекишечные поражения — от цистита до сепсиса; такие инфекции обычно возникают эндогенно, за счет штаммов, колонизирующих кишечник;
- диареегенные эшерихии, которые имеют экзогенное происхождение, не задерживаются в организме (транзиторные штаммы) и с этой точки зрения могут считаться патогенными разновидностями кишечной палочки.
Острые кишечные инфекции
Заражение кишечной палочкой происходит тотчас после рождения и завершается бессимптомной колонизацией толстого кишечника, где эшерихии доминируют среди аэробной (точнее факультативно-анаэробной) микрофлоры. Незаметно проходит и инфицирование большинством штаммов в дальнейшей жизни. Гораздо реже развивается острая (диарейная) патология кишечника — эшерихиозы или кишечная коли-инфекция. Диареегенность (т.е. способность поражать эпителий кишечника) зафиксирована у представителей многих О-групп, хотя внутри каждой из них не все штаммы обладают такой возможностью. О-антиген маркирует потенциальную болезнетворность, реализация которой требует дополнительных факторов.
Эшерихиозы относятся к антропонозам с фекально-оральным механизмом передачи. Исключение составляют энтерогеморрагические эшерихии, резервуаром которых служит, по-видимому, крупный рогатый скот. Другие диареегенные эшерехии тоже циркулируют среди животных, птиц и даже насекомых (тараканов), но это вряд ли имеет эпидемическое значение.
Ведущая роль принадлежит пищевому пути передачи, прежде всего через молоко и молочные продукты. Вторым по значению является водный путь. Контактно-бытовая передача (предметы обихода или ухода за ребенком, руки матерей, персонала детских учреждений и больничных стационаров) возможна при заражении грудных детей ЭПКП-штаммами, где ЭПКП — энтеропатогенные кишечные палочки (см. ниже).
Агрессивность диареегенных эшерихий невысока. Патогенетически значимая дозировка составляет 106—109 бактерий, что на несколько порядков больше, чем у сальмонелл брюшного тифа и шигелл (исключением являются энтерогеморрагические кишечные палочки). Восприимчивость сильно зависит от штаммовых особенностей и гораздо выше у детей младшего возраста, особенно первого года жизни. Приобретение возрастной устойчивости может быть связано с вакцинальным (протективным) эффектом эшерихий-комменсалов, располагающих общими антигенами с вирулентными штаммами, а также с субклиническим инфицированием диареегенными эшерихиями.
Инкубационный период обычно составляет 1—3 дня и никогда не превышает 7 дней. Главным источником инфекции служат больные (нередко со стертыми формами заболевания), выделяющие с фекалиями огромное количество возбудителя. Меньшее значение имеют реконвалесценты и бактерионосители. Это связано с тем, что носительство редко затягивается на время более 2—3 нед, а количество выделяемых бактерий настолько мало, что не представляет прямой угрозы. Опасным является загрязнение продуктов питания, в которых кишечная палочка находит подходящие условия для размножения. Этим объясняется эндемичное распространение эшерихиозов в жарких регионах с низким уровнем санитарной культуры. В промышленных странах умеренного пояса коли-инфекция встречается редко.
На основании патогенетических механизмов, серологических маркеров и эпидемиологических особенностей дифференцируют несколько основных разновидностей диареегенных эшерихий:
- энтеротоксигенные,
- энтеропатогенные,
- энтероинвазивные,
- энтерогеморрагические,
- энтероагрегирующие (энтероадгезивные).
Их общей особенностью является способность колонизировать кишечный эпителий, преодолевая (благодаря фимбриальным антигенам) элиминирующее противодействие перистальтики и конкуренцию со стороны нормальной микрофлоры. Генетика болезнетворности связана с двумя главными группами генов — плазмидами и хромосомными участками, получившими название островов патогенности. Некоторые признаки закодированы в транспозонах (например, термостабильный энтеротоксин) или в умеренных фагах (шигаподобные токсины энтерогеморрагических эшерихий).
Энтеротоксигенные кишечные палочки (ЭТКП) обнаружены среди представителей более 70 О-групп, чаще среди О6, О78, 0128 и О153 (в сумме они составляют более половины ЭТКП). Они продуцируют токсины, нарушающие баланс между секрецией и всасыванием жидкости эпителиальными клетками тонкого кишечника. Избыток воды и электролитов в просвете кишечника вызывает диарею.
Возможны различные формы заболевания — от легкой диареи до холероподобной интоксикации (малая холера). Лишь в редких случаях (чаще у недоношенных новорожденных) дегидратация требует неотложной помощи (регидратационной терапии). ЭТКП остаются главной причиной спорадических и эпидемичных детских диарей в развивающихся странах тропического и субтропического поясов, отвечая за 20—40% острых кишечных инфекций среди младенцев, находящихся на искусственном вскармливании. В экономически развитых странах ЭТКП заявляют о себе главным образом в виде диареи путешественников, которая наблюдается у туристов, посещающих неблагополучные по ЭТКП-инфекции регионы. Источником инфекции служит вода и пищевые продукты, загрязненные фекалиями, где возбудитель получает возможность для накопления. Обычно заболевания возникают в теплые и влажные месяцы, когда размножение эшерихий во внешней среде происходит особенно интенсивно.
Распространение бактерий ограничено поверхностью слизистой оболочки тонкого кишечника, энтероциты не инвазируются и структурно не повреждаются. Об этом, в частности, свидетельствуют отсутствие воспалительной реакции в стенке кишечника и водянистые испражнения без примеси слизи и крови. Такой тип диареи называют секреторным, противопоставляя инвазивному варианту, который наблюдается при деструкции кишечного эпителия. Секреторная диарея возникает вследствие гиперсекреции Na+, Сl— и воды каемчатыми клетками крипт при ослаблении альтернативной функции тонкого кишечника — всасывания электролитов и воды ворсинчатыми клетками слизистой оболочки.
Наиболее известны два эшерихиозных энтеротоксина — термолабильный (LT) и термостабильный (ST). Они закодированы в плазмидах — дискретных или общих. В последнем случае оба токсина синтезируются одновременно (около 5% штаммов).
LT-токсины образуют примерно 25% штаммов ЭТКП. По механизму действия (АДФ-рибозилтрансфераза) и даже по антигенным свойствам они похожи на холерогены — энтеротоксины холерного вибриона. После рецепции ганглиозидами энтероцитов LT-токсин (точнее, его А-субъединица) проникает в клетку и инактивирует (АДФ-рибозилирует) регуляторный белок, контролирующий активность аденилатциклазы. Диареегенный эффект связан с повышением внутриклеточного уровня циклического аденозинмонофосфата (рис. 2).

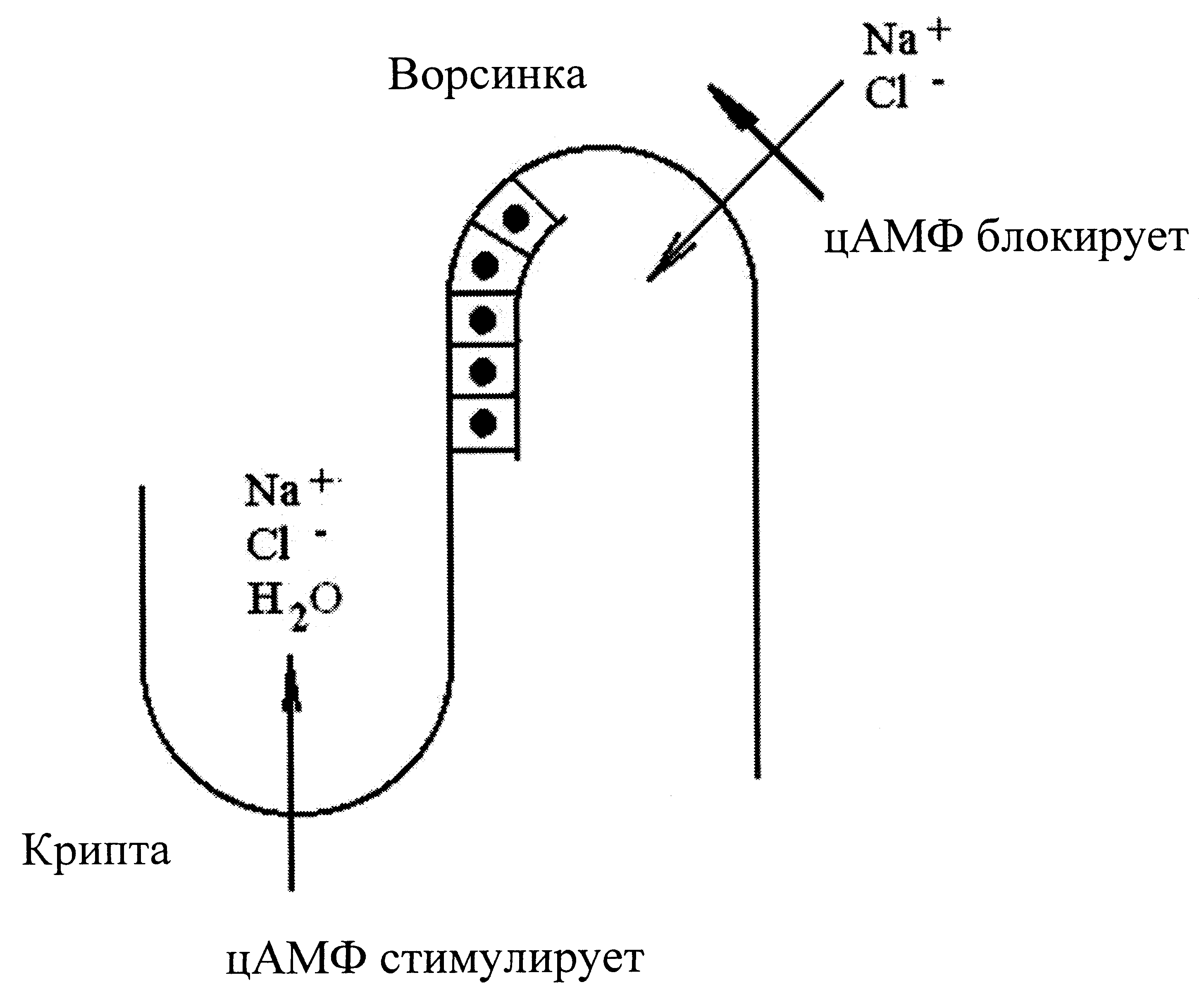
ST-токсин (его продуцируют около 70% штаммов ЭТКП) выглядит и действует иначе, хотя с патогенетической точки зрения делает «общее дело». Он состоит из двух низкомолекулярных пептидов (StA и StB), которые обладают слабой иммуногенностью и устойчивы к прогреванию, протеолитическим ферментам и кислотам. Токсин не проникает в клетку, действуя на рецепторы энтероцитов, связанные с мембранной гуанилатциклазой. Усиление синтеза циклического гуанозинмонофосфата нарушает внутриклеточный баланс по циклическим нуклеотидам, провоцируя гиперсекрецию воды и электролитов.
Токсинообразованию предшествует закрепление ЭТКП на энтероцитах, особенно проксимальных отделов тонкого кишечника. Это требует нестандартной атрибутики, которой лишены обычные кишечные палочки, населяющие толстый кишечник. Среди факторов, обеспечивающих избирательную колонизацию энтероцитов, лучше всего изучены CFA/I, CFA/II и CFА/IV (от англ. colonization factor antigens). Они экспрессированы примерно на 75% человеческих ЭТКП, примерно поровну распределены между ЭТКП-штаммами и представляют собой лектины, структурно оформленные в виде пилей/фимбрий (рис. 3).

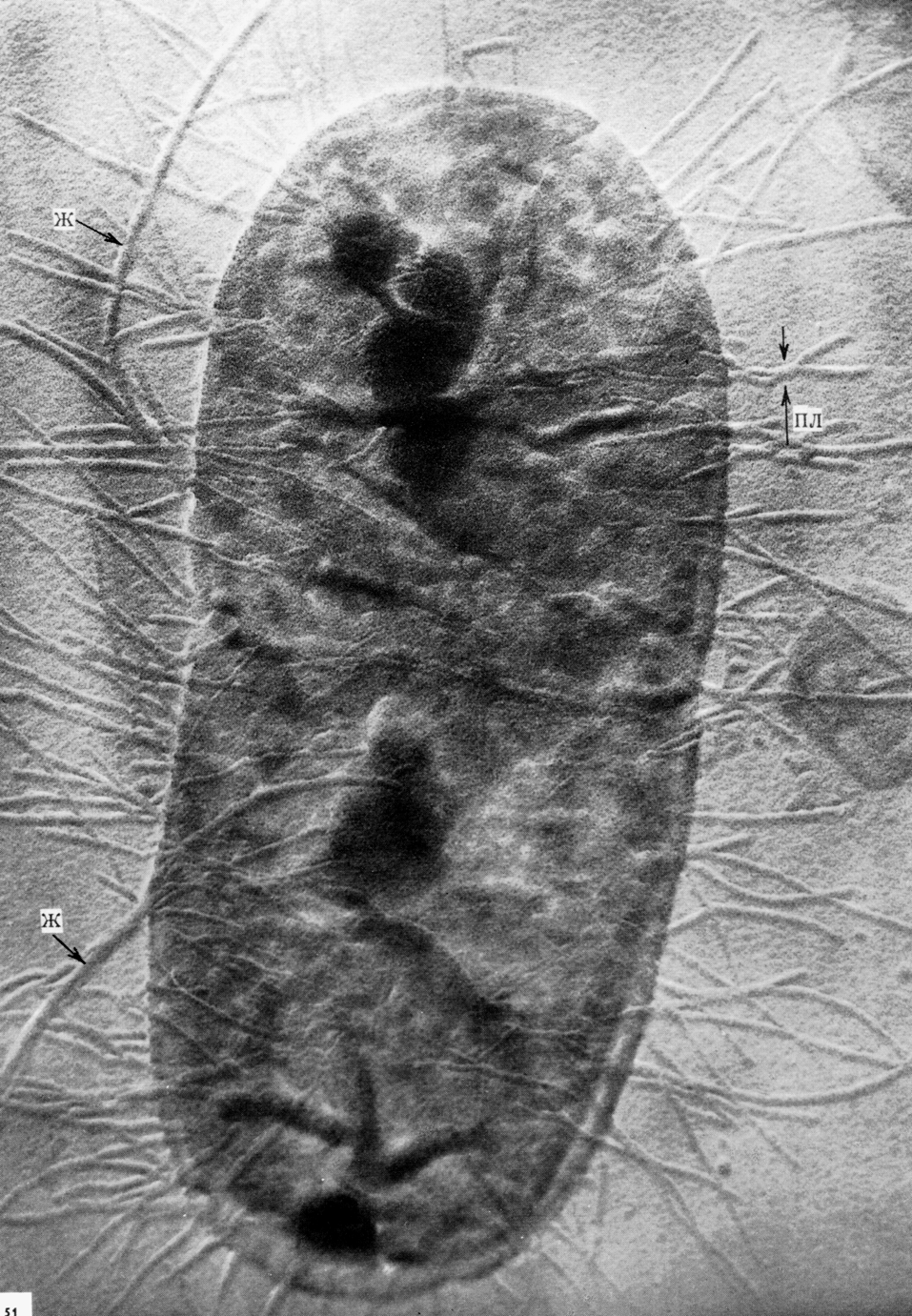
По одной из классификаций принято дифференцировать маннозочувствительные и маннозорезистентные адгезины эшерихий. Первые взаимодействуют с маннозными радикалами гликопротеинов и поэтому блокируются маннозой и ее производными, вторые — не реагируют с маннозой. CFA маннозорезистентны, что выделяет их из обширного арсенала фимбрий и нефимбриальных факторов, задействованных в банальной колонизации толстого кишечника. В отличие от них CFA воспринимаются теми же клеточными ганглиозидами (GM1), которые фиксируют LT-токсины. Заметим, что фимбрии ЭТКП животных (К88, К89, 987Р) тоже резистентны к маннозе, хотя и не идентичны адгезинам человеческих ЭТКП. Это объясняет причину антропонозности энтеротоксигенных коли-инфекций: ЭТКП животных не способны колонизировать тонкий кишечник человека, что, кстати, справедливо и для других категорий диареегенных эшерихий (см. ниже). Энтеротоксины ЭТКП человека и животных структурно тоже не идентичны, хотя и близки по механизму действия. Кстати, CFА/II и CFА/IV сами по себе неоднородны. Каждый из них состоит из трех антигенов — CS1, CS2, CS3 (CFA/II) и CS4, CS5, CS6 (CFA/IV) (от англ. cell surface). CFА/I структурно гомогенен.
CFA закодированы в плазмидах по соседству с генами энтеротоксинов, и лишь одновременная экспрессия cfa- и tox-генов обеспечивает ЭТКП-вирулентность. Без колонизирующих факторов энтеротоксины патогенетически инертны точно так же, как CFA-адгезины без токсигенности. Изучение факторов колонизации продолжается. К этому побуждают наблюдения за штаммами ЭТКП, лишенными CFA: они составляют около 20% выделяемых культур.
Энтеропатогенные кишечные палочки (ЭПКП). С них в 1950-х гг. стартовал современный этап в изучении диареегенных эшерихий. Этим объясняется неудачный универсализм термина (энтеропатогенность), который в равной мере можно распространить на все диареегенные эшерихии. ЭПКП, к которым относятся представители около 20 О-серогрупп (чаще О55, О111, О119, О127, О128), вызывают поражение тонкого кишечника у детей младшего возраста (до двух лет, обычно на первом году жизни). Ранее заболевание именовалось токсической диспепсией.
ЭПКП поражают тонкий кишечник. В основе диарейного синдрома лежит феномен сглаживания эпителиальных микроворсинок на фоне интимной адгезии между бактериями и эпителиальной плазматической мембраной. Это сопровождается образованием пьедесталоподобной структуры из-за аккумуляции филаментозного актина и других цитоскелетных элементов, таких как a-актинин, талин, эзрин и легкая цепь миозина (рис. 4). Драматичная потеря адсорбирующих ворсинок в зоне, прилегающей к месту адгезии ЭПКП, ведет к диарее за счет нарушения электролитного баланса и мальабсорбции.

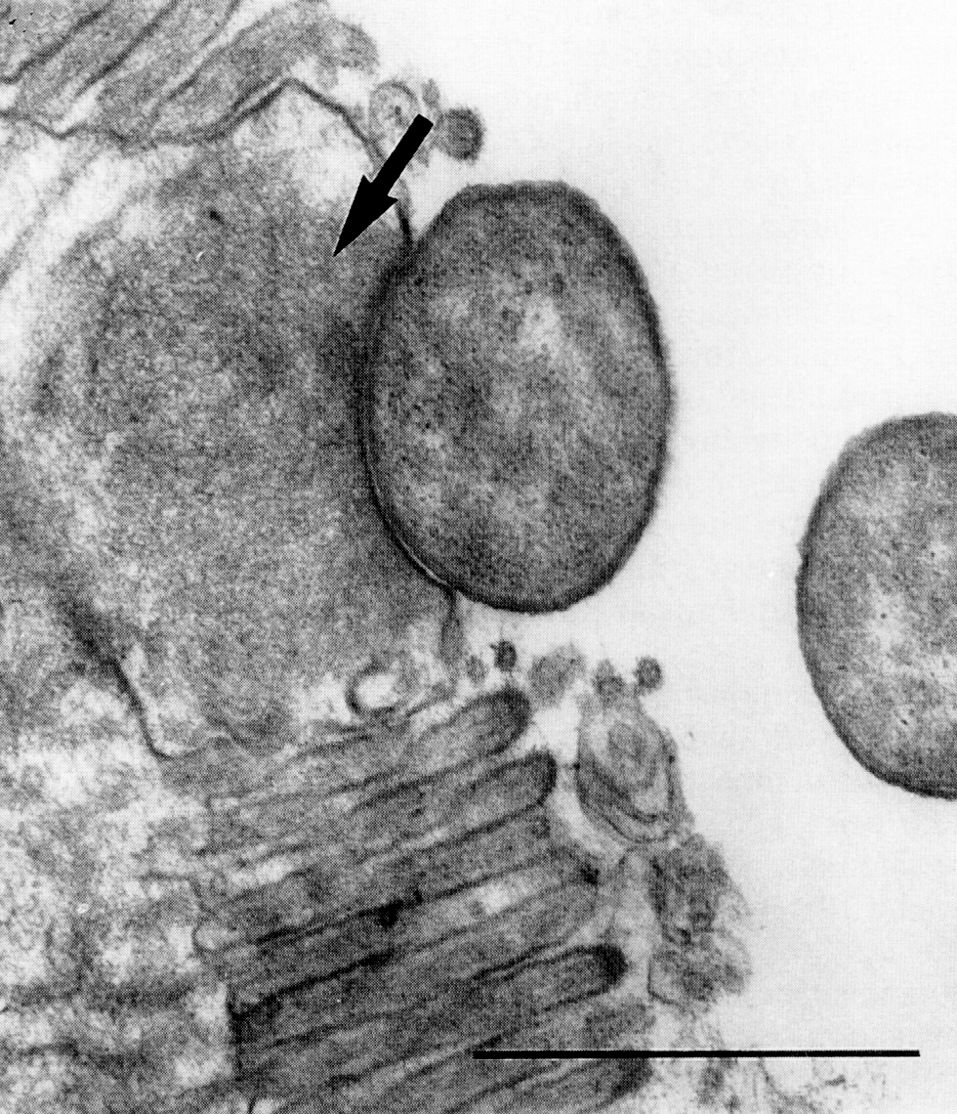
Расшифровка этого явления занимает одну из ключевых позиций в изучении патогенетических взаимоотношений между бактериями и клетками хозяина, затрагивая такие важные сюжеты, как природа адгезивных контактов, секреция III типа, функции эффекторных белков, перестройка внутриклеточного гомеостаза и пр. Патогенез ЭПКП-инфекции можно представить следующим образом. Попав в кишечник, ЭПКП начинают экспрессировать комплекс молекул, необходимых для закрепления бактерий на энтероцитах, и cинтеза белков, составляющих основу секреции типа III, — фимбрии (Bfp, от англ. bundle-forming pili), интимин, Tir (от англ. translocated intimin receptor), наружные белки (EspA, EspB, EspD). После адгезии ЭПКП впрыскивают в клетку эффекторные белки, которые активируют сигнальные каналы, вызывая изменения клеточного цитоскелета, выражающиеся в деполимеризации актина и потере микроворсинок. Заключительный этап возникает вслед за фосфорилирующей активацией Tir, включенного в плазматическую мембрану, и утратой первичных (Bfp- и EspA-зависимых) контактов между ЭПКП и энтероцитом. Интимин связывается с модифицированным Tir, приводя к интимной адгезии за счет образования массивного пьедестала из филаментозного актина и других компонентов цитоскелета в зоне бактериальной адгезии. Еще один эффектор, Мар (от англ. mitochondrial-associated protein), нацелен на митохондрии, производя разрыв мембранного потенциала. Нарушению интестинального барьера способствуют и другие факторы (EspF и др.), которые повреждают межклеточную интегративность, вызывая гибель клеток. Следует добавить еще один механизм ЭПКП-инфекции — воспалительный синдром, который сводится к трансмиграции нейтрофилов в зону поражения в ответ на цитокины (хемокины), секретируемые пораженными энтероцитами. Воспаление не очень значительно, хотя принципиально отличает ЭПКП от диареи секреторного типа, вызываемой ЭТКП.
Но это лишь общие представления. Имеется немало исключений, связанных с атипичными ЭПКП. Они требуют дополнительных исследований для правильного понимания механизмов болезни и разработки адекватных подходов к диагностике детских диарей.
Энтероинвазивные кишечные палочки (ЭИКП) вызывают синдром, практически не отличимый от шигеллезной дизентерии — диарею инвазивного типа (кровянистые испражнения с примесью слизи и лейкоцитов). Подобно шигеллам, ЭТКП проникают и размножаются в эпителиоцитах толстого кишечника и, повреждая их, индуцируют воспаление и изъязвление слизистой оболочки. Энтероинвазивность ограничена представителями нескольких О-групп: О28, О112, О124, О136, О143, О144, О152, О164. Они отвечают за небольшой процент всех случаев бактериальной дизентерии, значительно уступая шигеллам. Но этого достаточно, чтобы сделать проблему ощутимой в абсолютном исчислении, особенно при низком уровне санитарной культуры. ЭИКП обычно поражают детей — спорадически или в виде вспышек в организованных коллективах. Взрослые страдают редко — во время водных или пищевых вспышек.
Энтероинвазивность лучше всего отражает плотное родство между эшерихиями и шигеллами (гомология их ДНК превышает 90%). Это заставляет искать промежуточные формы, разделяющие данные таксоны энтеробактерий. Именно к таким гибридам относится большинство энтероинвазивных штаммов кишечной палочки. Многие из них были обнаружены благодаря своей атипичности, т.е. наличию признаков, общих с шигеллами: неподвижность, анаэрогенность (ферментация углеводов без газа), замедленная ферментация лактозы или полное отсутствие этого признака, родство по О-антигенам. Об их инвазивности судят по способности вызывать кератоконъюнктивит при закапывании бактериальной взвеси в конъюнктивальный мешок морской свинки или по цитопатическому эффекту в культурах эпителиальных клеток. Таксономические сомнения устраняют за счет расширения набора биохимических тестов. По ряду признаков (продукция лизиндекарбоксилазы, ферментация цитрата, утилизация ацетата натрия в качестве единственного источника углерода) такие культуры отличаются от шигелл и рассматриваются как атипичные эшерихии. Классические (т.е. быстро разлагающие лактозу) штаммы иногда тоже энтероинвазивны, хотя при внимательном анализе у них удается обнаружить признаки фенотипического подобия шигеллам (например, отсутствие лизиндекарбоксилазы).
Сходство с шигеллами распространяется и на механизмы, обеспечивающие инвазивность. Они зависят от генов, гомологичных генам вирулентности шигелл. Эффекторы инвазии точно не известны, но их гомология с факторами шигелл, доставляемых в энтероциты путем секреции III типа, несомненна. Инвазия начинается с пенетрации М-клеток кишечного эпителия и эволюционирует через латеральные контакты эпителиоцитов. Внутри клеток бактерии продуцируют ферменты, которые лизируют стенку фагосом, вызывая повреждение эпителиоцитов и распространение инфекции. Каким образом неподвижные ЭИКП становятся подвижными, можно судить по аналогии с шигеллами, которые обретают миграционную активность за счет внутриклеточного актина, прикрепляемого к одному из полюсов бактерий. В целом, подобно шигеллам, инвазивный фенотип складывается под влиянием хромосомных и плазмидных генов. ЭИКП и большинство серотипов шигелл имеют 140 MДa-плазмиду, Shigella sonnei — 120 MДa-плазмиду.
Справедливости ради заметим, что ЭИКП продуцируют один или несколько секреторных энтеротоксинов. Они отвечают за водную (секреторную) диарею, которой и заканчивается большинство случаев ЭИКП-инфекции. Только у единичных больных по-настоящему развивается дизентерийный синдром.
Энтерогеморрагические кишечные палочки (ЭГКП). С точки зрения вирулентности, это, пожалуй, наиболее оснащенная разновидность эшерихий. В формировании их болезнетворности участвуют гены острова патогенности, детерминирующие контактное взаимодействие (секрецию III типа) с мукозальным эпителием, гены умеренного фага, определяющие образование патогенетически значимого цитолизина, и плазмидные гены, которые кодируют дополнительные признаки вирулентности.
Большинство ЭГКП относится к серогруппе О157 (серотип О157:Н7), реже к О26, О111, О145. ЭГКП колонизируют толстый кишечник (особенно слепую кишку), вызывая геморрагический колит. В отличие от шигеллезных и ЭИКП-поражений кровянистые испражнения бедны лейкоцитами. Заболевание сопровождается общей интоксикацией (тошнота, рвота), а в самых тяжелых случаях (они наблюдаются у 10% детей до десяти лет) — внекишечными симптомами, к наиболее грозным из которых относится гемолитико-уремический синдром (гемолитическая анемия, тромбоцитопения, острая почечная недостаточность). Около 5% больных умирают, у 12—30% возникают тяжелые осложнения, включая почечную недостаточность, гипертонию и поражение центральной нервной системы. У взрослых инфекция протекает легко, не требуя специального лечения.
Инициирующая роль принадлежит агрессивной интиминзависимой адгезии, подобной той, что была описана выше для ЭПКП (полагают, что этого достаточно для провокации негеморрагической диареи). За ней следует продукция токсинов, которые оказывают местное действие (геморрагический колит) и вызывают системные эффекты (поражение почек, центральной нервной системы, внутрисосудистый гемолиз). Наибольшее значение имеют специфические цитолизины ЭГКП, называемые веротоксинами (по токсическому действию в культуре клеток линии Vero) или шигаподобными токсинами (подразумевая сходство с токсином Shigella dysenteriae). Это главный фактор вирулентности, который включает два серологически независимых токсина, Stx1 и Stx2, и множество подтипов Stx2. Токсины состоят из двух субкомпонентов, А-В. В-элемент связывается со специфическим гликолипидным рецептором клеток (глоботриозилцерамидом, Gb3/Gb4), обеспечивая внутриклеточное поступление А-субкомпонента. Благодаря блокаде белкового синтеза это ведет к прекращению метаболизма и гибели клетки. Наличие Gb3 (Gb4 для Stx2) является обязательным для действия шигаподобных токсинов, и именно этим объясняется их эффект на гломерулярный эндотелий почек и мелких сосудов, которые в больших количествах содержат такие рецепторы.
Шигаподобные токсины закодированы в умеренных бактериофагах, включенных в состав бактериальной хромосомы. Все штаммы О157:Н7 имеют плазмиду с мол. массой 60 МДa, в которой расположены гены для ряда других потенциальных факторов вирулентности, в частности, энтерогемолизинов. Однако их значение в болезнетворности ЭГКП подлежит уточнению. Не все ясно и с секрецией III типа: известны случаи гемолитико-уремического синдрома, когда выделялись штаммы О157:Н7 без генов, для интиминзависимой адгезии. Сложность патогенетического профиля ЭГКП не вызывает сомнений. Можно размышлять лишь над лидерством отдельных факторов и токсинов, не забывая о вероятности кооперативных эффектов.
В отличие от других коли-инфекций ЭГКП-эшерихиозы являются скорее всего зоонозами. Наиболее вероятным источником заражения считается крупный рогатый скот. Об этом свидетельствует ряд вспышек, зафиксированных в Канаде, США и Японии. Заражение происходит при употреблении в пищу мяса (после недостаточной термической обработки), сырого молока, воды. Имеют значение инфицированные овощи, фрукты и их дериваты. Впрочем, не все спешат отказываться и от антропогенности, предпочитая говорить об антропозоонозном характере инфекции. Передача возбудителя между людьми тоже возможна. Это стало понятным после того, как была установлена низкая инфицирующая доза ЭГКП: для клинически значимого заражения необходимо всего 100—200 клеток, что сопоставимо с шигеллезной инфекцией. Еще один парадокс: ЭГКП-эшерихиозом не болеют дети первого года жизни. Возможно, это тоже результат необычной для эшерихиозов эпидемиологии данной инфекции: новорожденные максимально защищены от контактов с зоонозными микробами.
Энтероагрегирующие кишечные палочки (ЭАКП). Дополнительным поводом к усложнению классификации диареегенных эшерихий явились морфологические особенности их адгезии в культурах эпителиальных клеток человека. Было замечено, что, в отличие от ЭПКП, для которых характерна локализованная адгезия с образованием поверхностных микроколоний, не-ЭПКП серотипы дают иную, диффузную морфологию взаимоотношений с Нер-2 клетками. На основании этого была выделена группа энтероагрегирующих кишечных палочек. Они склонны к аутоагглютинации и обусловливают фиксацию на клеточной мембране агрегатов в виде кирпичной кладки (рис. 5). Группа во многом неоднородна, включает патогенные и непатогенные клоны с одинаковым АА-фенотипом (АА, от англ. aggregative adherence).

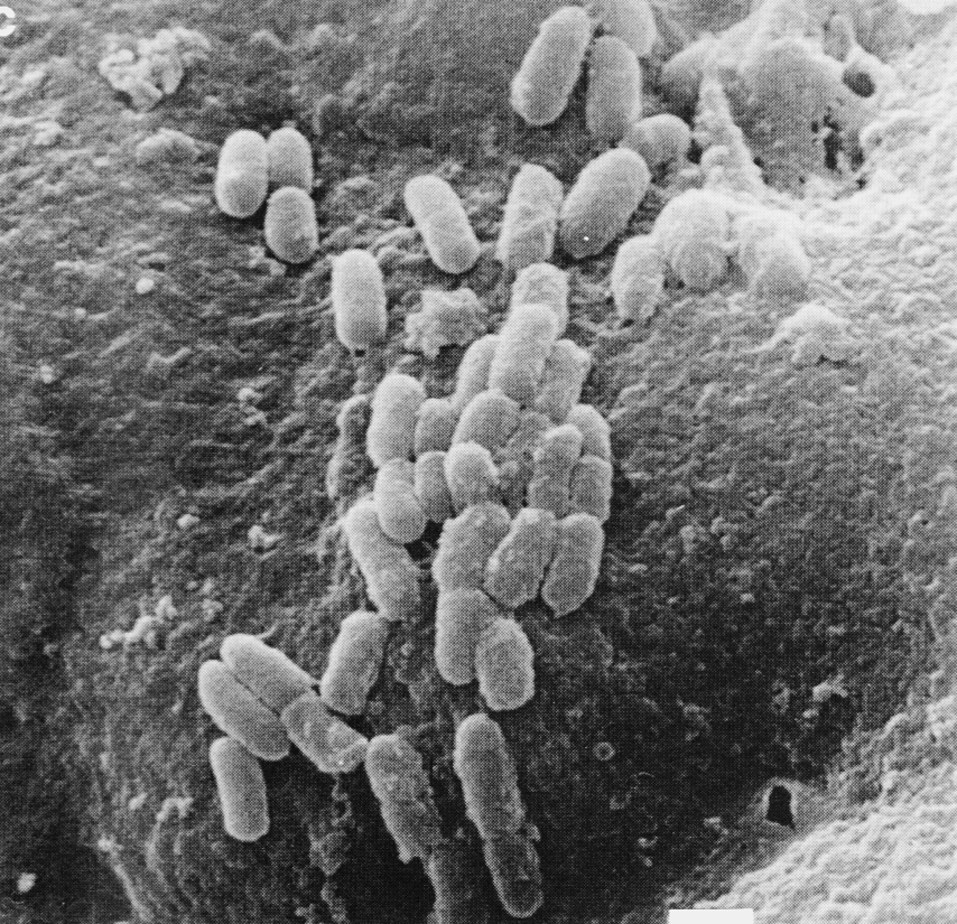
ЭАКП колонизируют тонкий и толстый кишечник, вызывая водную (секреторную) диарею, иногда осложненную воспалением слизистой оболочки и геморрагическим колитом. Болеют дети и взрослые, в основном из развивающихся стран (спорадическая эндемичная диарея, реже эпидемические вспышки). Нередко развивается персистентная диарея до 14 дней и более.
Структурная основа и патогенетическая значимость ЭАКП-вирулентности известны лишь приблизительно. Это отражает неоднородность группы, которая кроме клонов разной патогенности включает комменсальные штаммы. Описано по крайней мере три категории пилей/фимбрий (ААF/I, AAF/II, GVVPQ), которые связаны с агрегативной адгезией, но их значение в развитии болезни требует подтверждения. У ЭАПК обнаружен термостабильный энтеротоксин (EAST1), по-видимому, участвующий в патогенезе секреторной диареи. EAST1 часто сравнивают с StА энтеротоксигенных кишечных палочек, с которым он имеет ряд общих признаков. Но EAST1 экспрессируется почти 40% непатогенных E. coli и требуются дополнительные факторы, способствующие реализации его болезнетворности. Говорят, например, о цитотоксинах, один из которых подобен гемолизину уропатогенных E. coli. Делая «дырки» в мембране клеток, он обеспечивает повышение внутриклеточного уровня Са2+ и комплекса дестабилизирующих Са2+-зависимых реакций. Будущие исследования должны установить природу болезнетворного потенциала ЭАКП, вскрыв причину их гетерогенности. Уже сейчас описана группа энтеропатогенных диффузно агрегирующих E. coli, клонированы их первые адгезины и показано участие в патогенезе водных диарей у большинства инфицированных больных.
Внекишечные инфекции
Уроэпителиальные инфекции. Самым распространенным внекишечным эшерихиозом является пиогенное поражение мочевыделительной системы. Более 85% первичных пиогенных уропатий, так называемых бытовых инфекций (англ. community infections), регистрируемых у практически здоровых людей (чаще женщин) вне госпитальных условий, связано с кишечной палочкой. К этому предрасполагают анатомические особенности мочевыводящей системы и задержка мочи. Последнее обстоятельство характерно для позднего периода беременности, когда гормональное расслабление гладкой мускулатуры мочевыводящих путей сочетается с давлением увеличенной матки.
Бактерии, попадающие в мочевой пузырь из устья уретры, размножаются в застойной моче, вызывая воспаление (цистит). Поднимаясь по мочеточникам, инфекция захватывает почечные лоханки, поражая канальцы и паренхиму (пиелонефрит). Болезнь развивается остро, но иногда (особенно у детей) протекает хронически. В этом случае часто выявляется повышенная бактериурия (более 100 000 бактерий в 1 мл мочи), которая возникает без очевидных предрасполагающих факторов и не всегда сочетается с признаками цистита. Следует помнить, что умеренная бактериурия — нормальное явление, так как бактерии кишечника неизбежно попадают в устье уретры. До мочевого пузыря доходят немногие, и лишь единицы пробиваются выше. Приоритет эшерихий определяется тем, что они количественно доминируют среди кишечных бактерий, приспособленных для паразитирования в мочевыводящей системе. По неизвестным причинам уропатогенные штаммы кишечной палочки принадлежат к ограниченному числу О-групп: О1, О4, О6 и О18.
В основе уропатогенности лежит способность кишечной палочки к закреплению на эпителии мочевыводящих путей, которое выдерживает силу вымывающего тока жидкости. Концепция о монофакторной адгезии, на которую возлагались первые надежды, сменилась представлениями о поливалентности адгезивного процесса. Большинство адгезинов закодировано в сегментах хромосомы, именуемых островами патогенности. Здесь же находятся гены, детерминирующие другие признаки вирулентности (синтез сидерофоров, цитотоксинов и пр.). Известно около десятка островов патогенности (размером от 25 до 190 Кбайт), которые разбросаны по хромосоме и неодинаково представлены среди уропатогенных штаммов. Они определяют фенотипические особенности эшерихий, подтверждая обоснованность их патогенетической классификации.
Главной разновидностью пилей, специфичных для уропатогенных эшерихий, являются Р-фимбрии. Они отсутствуют у комменсальных и диареегенных штаммов и получили свое название из-за того, что чаще обнаруживаются у штаммов, ассоциированных с пиелонефритом. Р-фимбрии представлены многочисленными антигенными вариантами, которые связываются с одним и тем же гликосфинголипидным рецептором. Его специфика определяется глобобиозой (дисахарид галактозы: a-D-Gal-(1,4)-a-D-Gal), а заякоривание на клеточной мембране — церамидом.
Альтернативой служат фимбрии типа 1, экспрессируемые большинством штаммов кишечной палочки. Они реагируют с маннозными остатками гликопротеинов и поэтому не активны (не вызывают адгезии) в присутствии маннозы (маннозочувствительные фимбрии). Универсальность распространения на некоторое время выключила их из сферы интересов уропатологов. Отношение изменилось, когда было установлено, что при взаимодействии с маннозными радикалами уроплакинов (мукозальных гликопротеинов мочевого пузыря) фимбрии типа 1 инициируют внутриэпителиальную инвазию бактерий. Это может содействовать персистенции уропатогенных эшерихий, предрасполагая к рецидивам. В отличие от Р-фимбрий, которые являются обязательным (или по крайней мере важным) атрибутом пиелонефритогенности E. coli, фимбрии типа 1 чаще обнаруживаются у штаммов, вызывающих цистит. Не исключено, что фиксацию бактерий в разных отделах мочевыделительного тракта определяют разные адгезины или их активность неодинакова.
В патогенетически значимой адгезии уропатогенных эшерихий могут участвовать и другие адгезины. Установлена корреляция между уропатогенностью E. coli и F1C-фимбриями. Они были открыты более 20 лет назад, но лишь недавно стала известна молекулярная основа их взаимодействия с уроэпителиоцитами. F1C-фимбрии связываются с галактозилцерамидом и глоботриазилцерамидом, первый из которых экспрессируется эпителиальными клетками мочевого пузыря, мочеточников и почек, а второй обнаружен в почках.
Одним из факторов восходящей колонизации мочевыделительной системы может быть Dr-адгезин (Dr-гемагглютинин). Его рецепторы широко представлены в тканях, в том числе в почечной паренхиме. Рецепция возможна только у резус (Dr)-позитивных индивидов, что подчеркивает генетическую запрограммированность чувствительности к коли-инфекции. Впрочем, генетические особенности сглаживаются многофакторностью адгезии тем более, что тканевой тропизм других адгезинов более универсален. Например, S-фимбрии рецептируются сиалированными гликопротеинами (отсюда «S»), которые входят в состав базальных мембран эпителия и эндотелия и идентичны у всех людей.
Повреждение эпителия связано с пиогенностью уропатогенных эшерихий, т.е. со способностью инициировать и поддерживать нейтрофилзависимое воспаление. Факторы пиогенности точно не известны. В последнее время об этом чаще говорят с позиций контактной (адгезивной) болезнетворности E. coli. Вывод сводится к тому, что при взаимодействии с уропатогенными штаммами эпителиоциты отвечают быстрой продукцией нейтрофильных хемокинов, прежде всего ИЛ-8. Адгезивная пиогенность связана с двумя основными разновидностями фимбрий — Р и типа 1. Повышенный интерес к Р-пилям объясняется тем, что именно они служат отличительным признаком уропатогенных штаммов E. coli. Р-позитивные штаммы вызывают наиболее интенсивную трансэпителиальную миграцию нейтрофилов in vitro, а их интравезикальная инокуляция усиливает воспаление (выход нейтрофилов) в мочевом пузыре.
В моче нет опсонинов, и нейтрофилы «пробуксовывают», так как опсониннезависимое взаимодействие с фагоцитами не обеспечивает должного бактерицидного эффекта. Не следует забывать, что подобно всякой условной (оппортунистической) болезнетворности уропатогенность кишечной палочки — относительное понятие. При нарушениях нормального мочеиспускания пиурию могут спровоцировать штаммы, лишенные признанных факторов вирулентности. Это легко продемонстрировать на животных в опытах с механической задержкой мочи.
В наиболее общем виде механизм пиогенного поражения мочевыводящей системы выглядит следующим образом. Адгезия уропатогенных штаммов E. coli вызывает активацию эпителиальных клеток, побуждая их к секреции провоспалительных цитокинов (хемокинов). В этом участвует комплекс бактериальных адгезинов и клеточных рецепторов, причем каждая из гомологичных пар реализует сигнал не только через общие, но и собственные каналы. Эпителиальные хемокины (ИЛ-8 и др.) служат пусковым стимулом для трансэндотелиальной мобилизации нейтрофилов, которые выходят в ткани, а затем (преодолевая эпителиальный барьер) на мукозальную поверхность. По ходу этого маршрута они уничтожают бактерии, содействуя стерилизации уромукозального тракта, а также сами становятся источником медиаторов, которые поддерживают эволюцию воспалительного процесса, трансформируя его в хроническую или рецидивирующую болезнь. Этому способствуют нарушение мобилизационных реакций нейтрофилов и внутриэпителиальная инвазия бактерий, изолирующая их от эффекторов иммунитета.
Приоритет контактной (адгезивной) болезнетворности перед такими традиционными представлениями о механизмах вирулентности, как инвазия и цитодеструктивность, имеет серьезные основания. Попытки связать уропатогенность E. coli с повреждением эпителия не привели к однозначным результатам. Более того, один из известных цитотоксинов уропатогенных эшерихий, альфа-гемолизин, в нелитических дозировках инициирует секрецию провоспалительных цитокинов, т.е. ведет себя как модулин. Патогенетические функции другого экзотоксина, несмотря на грозное название — цитотоксический некротизирующий фактор 1, еще менее очевидны. Недавно описан вакуолизирующий цитотоксин, который вызывает повреждение эпителиоцитов почек и мочевого пузыря человека и, судя по экспериментам на мышах, принимает участие в патологии верхних отделов мочевыводящей системы. Однако токсины продуцируются не всеми штаммами. Напротив, пиогенная эпителиотропность — универсальный спутник уропатогенности, который хорошо вписывается в современные представления о микробных модулинах и реактивном (медиаторном) потенциале мукозальных эпителиоцитов.
Сепсис. Наиболее суровым проявлением системной инвазивности эшерихий служит гнойный менингит новорожденных (рис. 6).

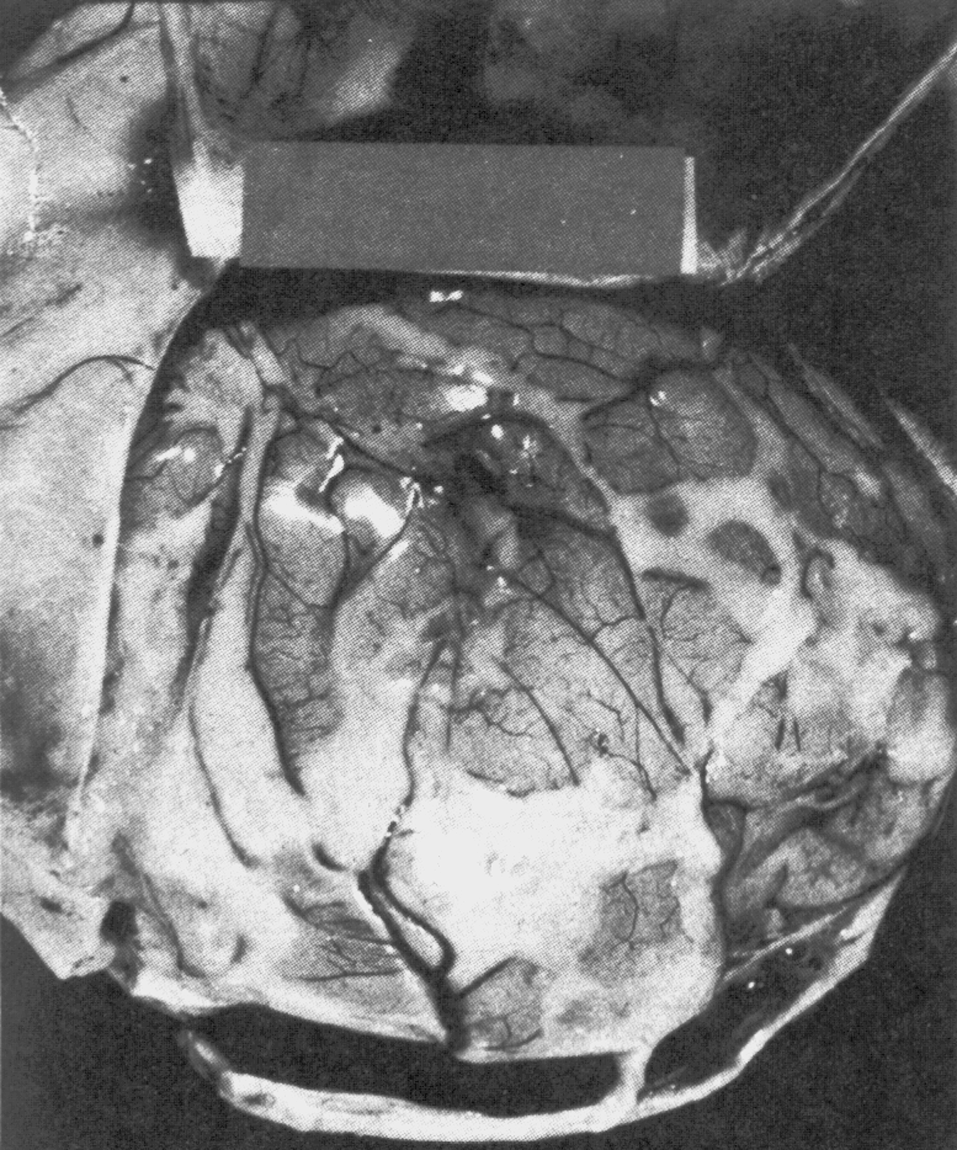
Он возникает в первые 30 дней жизни и наблюдается в 4 раза чаще у недоношенных. Смертность составляет 40—80%, и даже интенсивная антибактериальная терапия часто не спасает от тяжелых неврологических последствий. Маршрут бактерий складывается из пенетрации эпителиальных барьеров, проникновения в кровяное русло и далее (через фенестрированный эндотелий сосудистого сплетения головного мозга) в центральную нервную систему. Полагают, что этому способствует S-фимбриальная адгезия на сиалированных галактозидных радикалах ламинина базальной мембраны эндотелия или клеточносвязанного фибронектина. S-фимбрии закодированы в хромосоме и специфичны для серогруппы О18, доминирующей при данной патологии. Агрессивность усиливается и благодаря устойчивости в кровеносном русле. Она связана с особенностями К1-антигена, который обеспечивает резистентность к бактерицидному действию мембраноатакующего комплекса (С6—С9) комплемента. Лидерство принадлежит К1-позитивным штаммам, среди которых доминирует серовар О18:К1:Н7. Интересно (и, по-видимому, неслучайно), что К1 располагает общими эпитопами с полисахаридным антигеном менингококка группы В, одного из главных возбудителей менингита. Повышенная чувствительность новорожденных логично сочетается с дефектами иммунитета, присущими этому периоду онтогенеза. Антитела против О-антигена и капсульного полисахарида относятся к IgM, и, не получая их от матери (IgM не проходят через плаценту), новорожденные испытывают недостаток в этом наиболее мощном активаторе комплемента и комплементзависимого фагоцитоза. Следует учитывать и недоразвитие системы классического и альтернативного каскадов комплемента в раннем периоде новорожденности.
Эшерихиозный сепсис заявляет о себе и во «взрослой» медицине. Но здесь это, как правило, вторичные инфекции, которые осложняют врачебные манипуляции. Первое место здесь принадлежит колиформным бактериям, прежде всего кишечной палочке. Связанные с ней процессы носят откровенно оппортунистический характер, отражая потенциальную инвазивность эшерихий, для проявления которой требуется дискредитация иммунитетных ресурсов хозяина. Инфекция, как правило, возникает изнутри, благодаря активации собственной микрофлоры. Привязка к определенным серотипам необязательна или по крайней мере не столь очевидна. Вместе с тем патогенетические ресурсы различных штаммов неодинаковы, так как признаки, определяющие инвазивность, отчетливо клонированы. Речь идет о комплексе свойств, которые обеспечивают:
- устойчивость к механической элиминации (фимбриальные и нефимбриальные адгезины);
- резистентность к опсонофагоцитарным реакциям (факторы, интерферирующие с комплементом и нейтрофилами);
- растормаживание флогогенного потенциала фагоцитов и эпителиальных клеток (факторы, усиливающие продукцию цитокинов, хемотаксис и биотоксичность нейтрофилов);
- размножение бактерий при низком содержании свободного железа (сидерофоры типа энтерохелинов).
Несмотря на разнородность внекишечных изолятов, перечисленные свойства встречаются у них чаще, чем среди эшерихий, выделяемых из кишечника. Это означает, что подобно диареегенным разновидностям есть штаммы с повышенной «внекишечной проходимостью». Универсальным фактором общей интоксикации служит липополисахаридный эндотоксин, запускающий развитие септического синдрома.
Шигеллы
Не там вор ворует, где много, а там, где оплошно.
- Эпидемиология и клиника дизентерии (шигеллеза).
- Классификация шигелл.
- Адгезия, колонизация и внутриэпителиальная инвазия.
- Причины гибели макрофагов и колоноцитов.
- Токсин Шига и гемолитико-уремический синдром.
- Организация генов вирулентности: плазмиды и хромосома.
- Диагностика и лечение.
- Проблемы специфической профилактики.
Шигеллы (Shigella) — единственный род энтеробактерий, все представители которого патогенны для человека. Инфекции, которые они вызывают, являются антропонозами и в той или иной степени воспроизводятся только на обезьянах. Синонимом шигеллеза служит дизентерия — воспалительное заболевание толстого кишечника, протекающее с абдоминальными болями, интоксикацией, диареей с кровью, слизью и гноем. Это скорее клиническое, чем этиологическое понятие, так как симптомы дизентерии встречаются при ряде болезней, в частности протозойной (Entamoeba histolytica), вирусной (ротавирусы) и бактериальной (энтероинвазивные E. coli) природы. Среди возбудителей бациллярной дизентерии шигеллы занимают ведущее место.
Длительность инкубационного периода шигеллеза колеблется от 1 до 7 дней, чаще составляя 2—3 дня. Преимущественно поражается дистальный отдел толстой кишки с появлением поверхностных микроэрозий и кровоточащих язв. Наблюдается симптомокомплекс колита, когда слизистая оболочка воспалена и в ней присутствует выраженное количество нейтрофилов. Этому сопутствуют спастические боли в животе и выделение скудного бескалового содержимого со значительной примесью крови, слизи и гноя (нейтрофилов). Позывы (чаще бесплодные), сопровождаются мучительными тянущими болями в области прямой кишки — тенезмами. При выраженных формах шигеллеза наблюдается учащение стула до 20—30 раз и более в сутки. Дефекация, как правило, не приносит облегчения, а количество каловых масс, выделяемых больными, невелико — не превышает 0,5—1,0 л в сутки. К концу 1—2-х суток может развиться коллаптоидное состояние со смертельным исходом (по данным ВОЗ, в 1999 г. от шигеллеза погибло 1,1 млн. людей). Период разгара болезни колеблется от 1 до 9 дней, но полное выздоровление наступает через 3—6 нед. Примерно в 3% случаев наблюдается переход в хроническую форму, в 2% — формируется длительное бактерионосительство.
Нередко дизентерийный процесс протекает в легкой, стертой, субклинической или бессимптомной формах. Такие люди становятся источником инфекции, опасным для окружающих. Следует помнить, что Sh. sonnei (один из вариантов шигелл) интенсивно размножаются в пищевых продуктах, в частности в молоке. Способность некоторых из них выделять энтеротоксин ведет к его накоплению в пищевых субстратах. В сочетании с интенсивным размножением в тонком кишечнике это может вызвать клинический синдром по типу водной диареи, характерной для данного вида шигелл. Но главным остается колитический вариант болезни, который развивается вслед за пероральным (контактно-бытовым, водным или пищевым) поступлением бактерий в толстый кишечник. Страдают прежде всего дети до 10 лет в странах с низким санитарно-гигиеническим уровнем.
Впервые возбудитель бациллярной дизентерии был подробно изучен в 1898 г. японцем К. Шига (отсюда — шигеллы). Но раньше он был описан русским армейским врачом А.В. Григорьевым. Поэтому штаммы Sh. dysenteriae серотипа 1 называются Sh. grigorieva—shiga. Затем появились другие ферментативные варианты шигелл, каждый из которых вошел в историю под именем первооткрывателей (Sh. boydii, Sh. flexneri, Sh. sonnei и др.) или их называли в честь того места, где этот вариант был впервые выделен (Sh. newcastle). В конце концов это породило такую путаницу, что пришлось организовывать специальный международный комитет (1958 г.), который рекомендовал унифицировать классификацию шигелл.
Согласно разработанной систематике, род Shigella cостоит из четырех видов (или серогрупп), которые различаются по липополисахаридному О-антигену:
- Sh. dysenteriae (группа А);
- Sh. flexneri (группа В);
- Sh. boydii (группа С);
- Sh. sonnei (группа D).
На основании антигенных (эпитопных) различий О-антигена серогруппы делят на серологические типы, или варианты. Sh. dysenteriae объединяет 13 О-серотипов, включая наиболее опасный серотип — Sh. dysenteriae, тип 1. У Sh. boydii — 18 серотипов. Вид Sh. sonnei в этом отношении гомогенен, включает лишь один серотип, который, впрочем, существует в двух формах — I и II. Форма I кодируется плазмидой, спонтанная потеря которой ведет к утрате способности синтезировать антиген, характерный для первого серотипа. Поскольку он является одним из факторов, необходимых для внутриэпителиальной инвазии, форма I доминирует в процессе острой болезни. Штаммы формы II лишены вирулентности и наблюдаются в основном у носителей. Сложнее устроены Sh. flexneri. Они включают 6 серотипов, которые делятся на 11 подтипов. Кроме того, в этой группе есть штаммы, лишенные типоспецифических антигенов (х- и у-варианты). О-антигены липополисахаридных эндотоксинов являются единственным диагностически значимым антигеном шигелл. Н-антигенов у шигелл нет (они лишены жгутиков), а термолабильные К-антигены представляют интерес только потому, что от них приходится избавляться (кипятить), так как они мешают О-агглютинации.
Шигеллы являются грамотрицательными неспорообразующими палочками с типичными для энтеробактерий свойствами. Характерными (но не исключительными) отличиями шигелл служат неподвижность (отсутствие жгутиков), ферментация углеводов без газа (за исключением некоторых серотипов Sh. flexneri) и неспособность сбраживать лактозу (исключение составляют Sh. sonnei, медленно сбраживающие лактозу). Отметим еще одну особенность шигелл — отношение к манниту: все виды шигелл, за исключением Sh. dysenteriae, ферментируют маннит. Это служит важным диагностическим критерием в дифференциальной диагностике шигеллезных инфекций.
В нашей стране, как и в большинстве развитых стран Запада, преобладают возбудители двух видов шигелл — Sh. flexneri и Sh. sonnei, которые вызывают эндемичные формы заболевания. Sh. dysenteriae (особенно серотип 1) отвечают за смертельные эпидемии в развивающихся странах. Sh. boydii встречается только на Индийском полуострове. По тяжести развивающихся инфекций ряд шигелл выглядит следующим образом: Sh. dysenteriae — Sh. flexneri — Sh. boydii — Sh. sonnei.
Шигеллы относятся к инвазивным бактериям. Они обладают способностью колонизировать слизистую оболочку толстого кишечника и инвазировать ее в порядке факультативного внутриклеточного паразитизма, производя воспалительную деструкцию — ключевой элемент дизентерии. Причины этого, связанные с адгезией, инвазией и распространением шигелл в инфицированных клетках (энтероцитах), стали проясняться недавно, но их изучение еще далеко до своего завершения. Во-первых, не все благополучно с экспериментальными моделями шигеллеза. Бациллярная дизентерия — антропоноз и только на обезьянах может быть воспроизведена картина болезни, похожей на дизентерию человека. Это является серьезным недостатком, связанным с дороговизной опытов и трудностью получения солидного материала. Из других моделей отметим воспроизведение воспалительной реакции в кератоконъюнктивальной пробе на морских свинках и шигеллезного поражения респираторного тракта у мышей. Эти тесты воспроизводят флогогенный потенциал шигелл, но далеки от естественного заболевания человека. То же самое можно сказать и относительно инвитровых моделей в культурах поляризованных и неполяризованных клеток, которые позволяют судить о важных элементах патогенеза шигеллезной инфекции, но не воспроизводят всей картины дизентерии. Между тем в совокупности все эти модели все же позволяют судить о ключевых механизмах дизентерийной болезни у человека, и ниже мы попытаемся сформулировать основное содержание полученных выводов. Поскольку разные представители шигелл (Sh. flexneri, Sh. sonnei, Sh. boydii и Sh. dysenteriae) имеют сходные факторы вирулентности, они будут объединены вместе, хотя лучше всего изучен вид Sh. flexneri.
Шигеллы являются высокоинфекционными бактериями — для возбуждения болезни достаточно нескольких сот клеток, что значительно меньше, чем для диареегенных эшерихий (106—107) и S. typhi (105). Дойдя до толстого кишечника и прямой кишки, они колонизируют слизистую оболочку и вызывают ее разрушение, хотя природа этого явления полностью не раскрыта. Бактерии проникают в подслизистую оболочку, но обычно не идут дальше. Здесь после стимуляции и разрушения макрофагов происходит острая воспалительная реакция с привлечением нейтрофилов. Активированные нейтрофилы повреждают эпителиальный барьер, позволяя шигеллам возвратиться к эпителиальным клеткам, которые под их влиянием секретируют провоспалительные цитокины (хемокины). Последние, привлекая новые нейтрофилы, усиливают воспаление, приводящее к клинике дизентерии. Такова общая картина процесса. Теперь поговорим о том, что о нем известно.
Низкая инфекционная доза шигелл объясняется их относительно высокой устойчивостью (больше, чем у других энтеробактерий) к кислой среде желудка. Шигеллы чувствуют кислое окружение и адаптируются к нему благодаря регуляторным генам, экспрессия которых зависит от транскриптационного сигма-фактора (РНК-полимеразы, RpoS), который продуцируется в стационарной фазе роста. При этом у них снижаются инвазивные способности. Но как только бактерии достигают тонкого кишечника (щелочная или нейтральная среда) и вновь начинают размножаться (т.е. вступают в экспоненциальную фазу роста), их кислотная резистентность подавляется и восстанавливается инвазивный фенотип. Кислотная резистентность усиливается в условиях анаэробиоза, с которыми бактерии сталкиваются в толстом кишечнике. Поэтому шигеллы, экскретируемые с фекалиями, уже отличаются кислотоустойчивым фенотипом. Это готовит бактерии к заражению нового хозяина, когда они должны вновь пройти через кислую среду желудка.
Шигеллы не способны проникать в интестинальные эпителиоциты с их наружной (обращенной в полость кишечника) стороны. С этой целью они используют специализированные М-клетки эпителия, покрывающего лимфоидные фолликулы толстого кишечника. Эти клетки отличаются плохо развитой щеточной каймой, отсутствием гликокаликса и высокой эндоцитозной активностью. Они образуют пакеты, наполненные лимфоцитами, макрофагами и дендритными клетками, формируя контакты между кишечной микрофлорой и системой мукозального иммунитета. Полагают, что апикальная поверхность М-клеток несет рецепторы, воспринимающие шигеллы, и такой контакт запускает их секреторный аппарат III типа, в частности продукцию плазмидозависимых инвазивных антигенов IpaB, C, D, A (от англ. invasion plasmid antigens), которые транслоцируются в цитоплазму клетки-хозяина, принимая участие в реорганизации цитоскелета, нацеленной на инвазию шигелл.
Пройдя через М-клетки и оказавшись в lamina propria, шигеллы проникают в макрофаги. В отличие от сальмонелл, шигеллы не идут дальше, а возвращаются назад, в колоно(энтеро)циты. Это происходит после взаимодействия с макрофагами, требующего участия бактериальных инвазинов и клеточных рецепторов (в частности, aМb2-интегрина). Макрофаги высвобождают ИЛ-1 и ИЛ-18, которые запускают острое воспаление — приток нейтрофилов, характерный для шигеллеза. Это происходит благодаря IpaB, который секретируется аппаратом секреции III типа в цитоплазму макрофага и, связываясь с ИЛ-1b-конвертирующим ферментом (каспазой-1), способствует выделению цитокинов ИЛ-1 и ИЛ-18 из их предшественников. Через 1—2 ч после инфекции макрофаги погибают от апоптоза, хотя эта смерть отличается от классического апоптоза тем, что она не зависит от каспазы-3 и каспазы-11, а также не регулируется антиапоптозными белками семейства Bcl-2 (см. «Апоптоз»). В этой модели каспазу-1 следует рассматривать как компонент врожденного иммунного ответа, который использует апоптозный путь для выделения зрелых цитокинов, задействованных в остром воспалении.
Ключевым моментом дизентерии является способность шигелл инвазировать эпителий толстой кишки. В отличие от других инвазивных патогенов, шигеллы обладают слабой способностью связываться с клетками. В культурах клеток обнаружено взаимодействие наружных белков (инвазинов) шигелл с a5b1-интегрином (рецептор фибронектина) и/или CD44 (рецептор гиалуроновой кислоты). Это может индуцировать контактную реакцию аппарата секреции III типа, когда клетка получает от бактерий комплекс эффекторных факторов, приводящих к реорганизации цитоскелета и превращающих эпителиальную клетку в фагоцит, который поглощает шигеллы. Но эпителиальные клетки поляризованы, и шигеллы могут инвазировать их лишь с базолатеральной стороны, где имеются данные рецепторы. Дело решают нейтрофилы, спровоцированные к выходу в зону реакции цитокинами активированных макрофагов. Они мигрируют между клетками, открывая путь для бактериальной адгезии, внутрицитозольной секреции эффекторных белков бактерий, а затем и их инвазии в область базолатерального мембранного домена. Здесь сочетаются плюсы и минусы воспалительных реакций. С одной стороны, привлечение нейтрофилов выгодно бактериям, содействуя развитию инфекции. С другой — нейтрофилы эффективно борются с шигеллами в рамках врожденного иммунитета. Нейтрофилы, которые первоначально участвуют в повреждении тканей, в конце концов играют роль санирующего фактора.


Оказавшись внутри клеточной вакуоли, шигеллы в течение нескольких минут разрывают ее и выходят в цитоплазму. Бактерии получают здесь обильный питательный материал, быстро размножаются и распространяются по клетке и межклеточно через образующиеся пальцеобразные выросты на клеточной поверхности. Фагоцитоз этих выростов соседними клетками (в этом важную роль играет межклеточный Е-кадхерин) и лизис клеточных мембран позволяет шигеллам достичь цитоплазмы новых клеток, не выходя во внеклеточное пространство (см. рисунок). Для выхода из фагоцитарной вакуоли и распространения между клетками нужны IpaB и IpaC. Способность шигелл инвазировать эпителиоциты, выходить из фагосомы и подвергаться межклеточному распространению является ведущим фактором развития дизентерии. Потеря у мутантов одного из этих признаков резко снижает их способность к развитию воспаления.
Вопрос о том, каким образом шигеллы, лишенные жгутиков и активной подвижности, получают возможность для внутриклеточного и межклеточного передвижения, решился неожиданно. Оказалось, что бактерии после прорыва вакуоли, используя униполярную локализацию своего белка IcsA (от англ. intercellular spread), аккумулируют на одном из полюсов хвост из коротких пучков актина, который в качестве псевдожгутика проталкивает бактерии в соседние клетки. В лизисе выростов и высвобождении бактерий участвует протеин IcsB шигелл.
Отвечая на бактериальную инфекцию, колоноциты подвергаются активации. В этом, в частности, участвует липополисахаридный эндотоксин шигелл. Связываясь с тольподобными рецепторами (TLR4) на базолатеральной поверхности клеток, а также с внутриклеточными рецепторами Nod1 и Nod2, этот токсин активирует систему NF-kB (в меньшей степени АР-1), приводя к секреции цитокинов (хемокинов), в частности ИЛ-8 — мощного хемоаттрактанта нейтрофилов (см. «Воспаление и инфекционный процесс»). Это усиливает острое воспаление. Нейтрофилы выделяют множество биотоксических факторов, которые действуют не только на бактерии, но и на хозяина, вызывая расширение зоны местного поражения и системные реакции. Гибель нейтрофилов также зависит от бактериального аппарата секреции III типа, в частности белков IpaB и IpaC. Но шигеллы способны индуцировать и самостоятельную гибель колоноцитов. Их размножение ведет к прекращению синтеза белка и в конце концов к смерти клетки. Это скорее всего связано с утилизацией АТФ и нарушением функций митохондрий. Последнее обстоятельство может приводить к гибели клетки от апоптоза. Подавление синтеза белка вряд ли связано с действием экзотоксина Шига (см. ниже): атоксигенные мутанты убивают энтероциты так же, как дикие штаммы.
Вопрос о патогенетическом значении токсина Шига требует специального комментария. Он продуцируется штаммами только одного вида шигелл — Sh. dysenteriae серотипа 1. Подобно холерному энтеротоксину, дизентерийный токсин состоит из одной А-субъединицы (она отвечает за энзиматическую активность) и пяти В-субъединиц (рецепторная активность), которые закодированы на хромосоме в виде оперона так же, как А- и В-субъединицы холерного токсина. Механизм действия токсина Шига похож на действия других А—В-токсинов. Высвобождаясь при лизисе бактерий, он связывается с поверхностным гликолипидом (Gb3) клеток, субъединица А отщепляется и транслоцируется внутрь клетки. Действуя высокоизбирательно, А-субъединица инактивирует 60S рибосомы клетки, обрывая синтез белка.
Токсин Шига обладает рядом биологических эффектов, что делает Sh. dysenteriae серотипа 1 наиболее агрессивным агентом. Токсин Шига обладает активностью энтеротоксина, провоцируя выход жидкости в изолированную петлю кишечника, ведет себя как нейротоксин, вызывая параличи при введении мышам и кроликам, и, наконец, является сильным цитотоксином при добавлении в культуры клеток. Это делает понятной вирулентность токсинпродуцирующих штаммов шигелл в клинических наблюдениях и в опытах на животных (обезьянах). Но все-таки следует учесть, что шигеллы (например, Sh. flexneri), лишенные генов токсина Шига, вызывают тяжелое заболевание с серьезным поражением кишечника, которому сопутствует кровавая диарея. Поэтому допуская участие токсина Шига в поражении слизистой оболочки кишечника, необходимо искать для него другие патогенетически значимые мишени в инфицированном организме. Одной из них являются почечные капилляры (гломерулы), поскольку токсин Шига нацелен прежде всего на эндотелиальные клетки (кровавый понос — повреждение кровеносных сосудов в толстом кишечнике, неврологические осложнения — в центральной нервной системе). Поражение сосудов может отвечать и за острую почечную недостаточность (гемолитико-уремический синдром), которая развивается у детей через несколько дней после начала дизентерии и может быть смертельной. О сходном действии шигаподобного токсина энтерогеморрагических эшерихий уже говорилось в одном из очерков (см. «Эшерихии»).
Многие гены, необходимые для проявления вирулентности шигелл, локализованы на гигантских плазмидах. Лучше всего изучена 220-килобайтовая плазмида Sh. flexneri субтипа 2а, хотя и другие виды шигелл также располагают плазмидами подобного типа. Сходные плазмиды обнаружены и у некоторых штаммов E. coli (см. «Эшерихии»), что говорит о подвижности вирулентных плазмид среди энтеробактерий. Хотя на сегодня идентифицирована лишь часть генов вирулентности шигелл, можно сформулировать ряд общих закономерностей вклада в вирулентность плазмидных и хромосомных генов. Плазмида содержит примерно 20 генов, кодирующих аппарат секреции III типа, через который шигеллы запускают в клетки не менее 25 белков. Большинство структурных генов, необходимых для адгезии, инвазии и межклеточного распространения, локализованы на вирулентной плазмиде. Хромосомные гены тоже участвуют в инвазии шигелл, но это скорее регуляторные, чем структурные сигналы. Хромосома отвечает за внутриклеточный рост и размножение шигелл, а также за продукцию липополисахаридного эндотоксина и токсина Шига (Sh. dysenteriae). В целом можно говорить о том, что гены, принимающие участие на более поздних стадиях шигеллеза, имеют хромосомную природу; бесплазмидные мутанты лишены вирулентности. С плазмидами, несущими транспозоны антибиотикорезистентности, связана приобретенная устойчивость к антибиотикам (см. «Антибиотики»), которая сыграла трагическую роль в эпидемических вспышках дизентерии в Центральной Америке и Мексике в конце 1960-х — начале 1970-х гг.
Ведущим в диагностике шигеллеза остается культуральный метод. Обследованию подвергают испражнения больного. Так как шигеллы быстро гибнут в фекалиях, посев делают тотчас на агар либо на селенитовую транспортную среду (задерживает рост кишечной палочки и других комменсалов) для сохранения жизнеспособности бактерий. Исследуют лактозонегативные колонии, проверяя их ферментативные (безгазовая ферментация глюкозы, отсутствие ферментации сахарозы, образования сероводорода, индола, уреазной активности, подвижности) и антигенные свойства. И все-таки, несмотря на такие предосторожности, чувствительность метода варьирует от 20 до 80%. Для подтверждения используют серодиагностику — реакцию непрямой гемагглютинации эритроцитов, покрытых О-антигенами Sh. flexneri и Sh. sonnei. Учитывают минимальный диагностический титр антител (1:200) либо их нарастание в ходе болезни. Метод имеет ретроспективное значение: положительный результат может быть получен в конце первой недели заболевания. Разработаны экспресс-методы диагностики шигеллеза (иммунофлюоресценция мазков из фекалий, иммуноферментное выявление специфических антигенов, полимеразная цепная реакция для определения шигеллезных генов и др.). Они лишь дополняют, но не заменяют культурального анализа. То же самое можно сказать и о присутствии в стуле лейкоцитов. Их наличие говорит лишь об инвазивной диарее и может быть полезным диагностическим индикатором. Для клиники заключение о выделении определенного вида (серогруппы) шигелл является достаточным. Но для эпидемиологических целей культуры требуется изучить глубже, чтобы отдифференцировать штаммы одного вида и даже одного серотипа. С целью определения эпидемиологических маркеров применяются колицинотипирование (особенно для Sh. sonnei), фаготипирование, изучение плазмидного профиля, анализ полиморфизма рестрикционных фрагментов ДНК, пульсовый гель-электрофорез, риботипирование. С помощью этих методов удается установить тонкие свойства выделенных культур, сделав заключение об их происхождении в каждом конкретном случае.
Наиболее эффективными из современных антишигеллезных препаратов являются фторхинолоны. Следует, однако, помнить, что целесообразность их назначения определяется тяжестью заболевания. То же самое можно сказать и о назначении регидратационной и дезинтоксикационной терапии.
Специфической вакцинации не проводится, так как нет лицензированных вакцин, разрешенных к применению на человеке. Главной задачей является получение живой аттенуированной вакцины, которая при пероральном приеме будет создавать местный иммунитет кишечника. В этой связи возрос интерес к возможности использования парциальных антигенов шигелл для получения антиадгезивных и антиинвазивных антител. Такие препараты проходят испытания в экспериментальных системах, но окончательное заключение может дать только проверка на людях, так как ни одна из моделей на животных не воспроизводит дизентерии у человека.
Сальмонеллы
- Классификация сальмонелл.
- Кризис таксономики и его ликвидация.
- Эпидемиология, клиника и патогенез сальмонеллезов.
- Брюшной тиф: местная инфекция, переходящая на системный уровень.
- Исходы сальмонеллезных инфекций.
- Микробиологическая диагностика брюшного тифа.
- Специфическая профилактика брюшного тифа и возможности этиотропного лечения.
Бактерии получили название в честь американского микробиолога Д. Сэлмона, выделившего в 1885 г. одного из первых представителей рода Salmonella (Salmonella cholerae-suis) во время эпизоотии холеры свиней. Сальмонеллы — типичные представители семейства Enterobacteriaceae, т.е. грамотрицательные неспороносные палочки (средний размер 1—3 мкм), как правило, подвижные, неприхотливые, быстро ферментирующие глюкозу (обычно с образованием газа, исключение — S. typhi), восстанавливающие нитраты до нитритов, не содержащие оксидазы, каталазопозитивные, факультативные анаэробы. Наиболее характерными родовыми признаками является то, что они не ферментируют лактозу и сахарозу, не утилизируют желатины, не образуют индола, лишены уреазной активности, не продуцируют ацетилметилкарбинола (и соответственно дают положительную реакцию с метилрот), утилизируют цитрат аммония, образуют сероводород.
Следующий уровень в классификации сальмонелл выделяют на основе серологических признаков, т.е. на изучении антигенной структуры штаммов (схема Кауфмана—Уайта). Как и у остальных энтеробактерий, главным служит выявление О-антигена — полисахаридного компонента эндотоксина. Это неоднородный комплекс, состоящий из нескольких эпитопов. По О-антигенам сальмонеллы разделены на группы — А, В, С, D, E и т.д. (всего более 60 вариантов), причем каждая О-группа характеризуется главным (он обычно стоит на первом месте) группоспецифическим О-антигеном. Кроме того, есть О-антигены, которые, не являясь группоспецифическими, определяются в составе сальмонелл, принадлежащих к разным группам. Более 98% культур сальмонелл, выделяемых у больных и носителей, относятся к пяти первым О-группам — А, В, С, Д, Е, поэтому на практике пользуются сокращенной схемой Кауфмана—Уайта.
Сальмонеллы в подавляющем большинстве являются подвижными бактериями, т.е. у них есть жгутики и соответственно флагеллярные (жгутиковые) Н-антигены. Подобно О-антигену, Н-антиген устроен сложно и имеет различные серологические варианты, по которым можно дифференцировать штаммы внутри О-групп. Практически различают два основных варианта, которые получили название «фаза». Различают антигены 1-й и 2-й фаз. Первые обозначаются маленькими латинскими буквами — специфическая фаза. Они высокоиндивидуальны в пределах О-группы, позволяя отличать серотипы (серовары) штаммов. Антигены 2-й фазы (их обозначают через двоеточие арабскими цифрами) встречаются у разных серотипов, т.е. являются неспецифичными. Некоторые из сальмонелл имеют только один, типоспецифический Н-антиген — их называют монофазными; другие штаммы являются двухфазными.
Определение антигенной структуры сальмонелл (как и других энтеробактерий) проводят в реакции агглютинации на стекле. Делают это в три этапа. Во-первых, исследуют реакцию с поливалентной О-сывороткой к группам А, В, С, D, Е. В случае положительного результата проводят реакцию с монорецепторными О-сыворотками к каждой группе отдельно. Затем испытывают культуру в реакции агглютинации с монорецепторными Н-сыворотками, вначале 1-й, потом 2-й фазы. Полная антигенная формула будет выглядеть, например, следующим образом — (1),4,5,12:i:1,2 (для группы В, серотипа Salmonella typhimurium). Группоспецифическим антигеном является О-антиген под номером 4 (четыре).
Затруднение вызывает наличие у большинства штаммов S. typhi термолабильного поверхностного полисахаридного антигена, называемого Vi. Это разновидность К-антигена, который получил свое название от слова «вирулентность», потому что с ним связывали болезнетворность возбудителя брюшного тифа. Позже оказалось, что это не совсем так, так как Vi-антиген может, например, обнаруживаться у относительно безобидных эшерихий. Располагаясь более поверхностно, он закрывает О-антиген, препятствуя агглютинации О-антителами. Чтобы добиться положительного результата (т.е. разрушить Vi-антиген), такой штамм надо прокипятить в течение 10 мин. Поэтому в антигенной формуле S. typhi указывают наличие Vi-антигена — 9,12 (Vi):d: — (группа D, монофазный вариант).
Впрочем, присутствие Vi-антигена характеризует важную особенность штаммов — их потенциальную чувствительность к Vi-фагам, которые рецептируются этим антигеном, а потому взаимодействуют только с Vi-позитивными штаммами сальмонелл. Это используется на практике в качестве эпидмаркера для более глубокой идентификации S. typhi. Cреди всех сальмонелл Vi-антиген обнаружен в основном у S. typhi.
Классификация сальмонелл усложнялась по мере их открытия. Это стало особенно заметно начиная с 1920-х гг., после обнаружения диагностических антигенов сальмонелл. Развился таксономический кризис. К нему привела ситуация, сложившаяся вследствие традиции присваивать большинству серотипов сальмонелл видовые эпитеты, в то время как эшерихии и прочие энтеробактерии были лишены их. Никто не мог предположить, что к концу ХХ века станет известно более 2000 близко родственных типов бактерий, каждый из которых будет иметь индивидуальную серологическую характеристику. Учитывая важность антигенных свойств сальмонелл, было решено упростить их классификацию. Сегодня выделяют единственный вид — Salmonella enterica, внутри которого дифференцируют О-группы, состоящие из множества Н-сероваров (Н-серотипов), о которых говорилось выше. Для того, чтобы не запутаться и не потерять связь с историей микробиологии, за каждым «старым» сероваром сохранено видовое (точнее псевдовидовое) название. Поэтому сегодня вместо S. typhi надо говорить S. enterica, серовар Typhi; S. typhimurium надлежит называть S. enterica, серовар Typhimurium и т.д. Вновь открытым серотипам рекомендовано не придумывать новых названий. Все логично, но эти указания пока не прижились, и практики продолжают использовать привычные термины. Мы тоже будем применять их в настоящем очерке.
Сальмонеллез
Болезни, вызываемые сальмонеллами у человека, делятся на две категории — брюшной тиф/паратифы и сальмонеллезы. Наиболее распространены сальмонеллезы. Обычно это диарейные поражения тонкого кишечника (энтериты), которые развиваются через 6—24 ч после приема зараженной пищи или воды и заканчиваются самоизлечением в течение недели. Первоначальные симптомы включают тошноту и рвоту, которые продолжаются несколько часов. Затем следуют абдоминальная боль, диарея, иногда температура. Степень боли и диареи широко колеблется — от легких ощущений до боли, симулирующей аппендицит, и тяжелой (даже кровавой) диареи. Особенно тяжело болеют дети до 10 лет. После исчезновения симптомов больной (реконвалесцент) продолжает выделять бактерии до 3 мес; в 1—3% случаев возникает хроническое носительство — выделение сальмонелл более года. С этим связаны случаи госпитальной инфекции — системной септикопиемии, которой чаще болеют дети, иммунокомпрометированные больные и пожилые люди.
В принципе любой из серотипов сальмонелл способен спровоцировать болезнь. Обычно это наблюдается с S. typhimurium, S. enteridis, S. anatum, S. derby, S. london, S. panama, S. newport и еще примерно с 40 серотипами сальмонелл серогрупп В, С, D, E. Они встречаются в кишечнике домашней птицы и сельскохозяйственных животных, как правило, безвредны для них, почти неизбежно попадают в пищевые продукты (яйца, мясо, молоко и др.) и воду и становятся опасными для человека. Работники сельского хозяйства и пищевой индустрии принимают все меры, чтобы снизить уровень заражения, но, по-видимому, никогда не будет пищи, свободной от сальмонелл. Единственное, что можно сделать — не употреблять залежалую провизию и следить за правильным приготовлением продуктов. Опасность представляет прием более 105 бактерий, а это означает, что сальмонеллы должны накопиться в пище, прежде чем она будет представлять реальную опасность.
Было и до сих пор ведется немало споров о том, почему и какие факторы сальмонелл являются вирулентными для человека. Оставляя в стороне риторический вопрос о cальмонеллезных энтеротоксинах, остановимся на том, что известно о S. typhimurium, наиболее изученном серотипе сальмонелл. Его название отражает характер болезнетворности для мышей: S. typhimurium при пероральном введении вызывает у них не энтерит, а смертельную системную инвазию, напоминающую брюшной тиф у человека.
Сальмонеллы должны пройти сквозь пищеварительный тракт, чтобы оказаться в зоне, оптимальной для своей первичной колонизации — дистальном отделе подвздошной кишки. Это ведет к большим потерям бактерий, так как сальмонеллы значительно чувствительнее к кислой среде желудка, чем, например, шигеллы. Клинически значимая доза S. typhimurium при нормальной кислотности желудка составляет 10—100 млн. бактериальных клеток, в то время как та же дозировка в смеси с нейтрализующим кислоту бикарбонатом натрия снижается в 10—100 раз.
Сальмонеллы располагают генами, среди которых следует отметить двухкомпонентную систему PhoP/PhoQ. Это глобальный регулятор вирулентности, который улавливает в окружающей среде различные сигналы (в том числе рН), приводя к транскрипционному включению или выключению более 40 генов. Динамическое изменение экспрессии этих и других генов позволяет бактериям лучше адаптироваться к меняющимся условиям внешней среды, проявляя свою вирулентность.
Сальмонеллы прикрепляются к энтероцитам с помощью комплекса хромосомных и плазмидных адгезинов, оформленных в виде пилей, и, подобно шигеллам, пенетрируют мукозальный барьер. Нельзя с уверенностью сказать, какие структуры при этом вовлекаются, так как все типы эпителиальных клеток чувствительны к инвазии сальмонелл. Тем не менее все больше данных говорит о том, что местом первичной колонизации служат М-клетки, покрывающие лимфоидные агрегаты. Ангажируя этот маршрут, сальмонеллы проходят к макрофагам, располагающимся под ними. Феномен связан с продуктами инвазивных генов (invA-E), которые внедряются в клетку при помощи аппарата контактной секреции III типа. Расположенные в острове патогенности (SPI-1), они влияют на цитоскелет хозяйской клетки, вызывая эндоцитоз бактерий путем раффления (англ. ruffling — волнение, рябь) ее плазматической мембраны. Это указывает на цитоскелетную перестройку, приводящую к формированию мембраносвязанной везикулы (рис. 1). Проникновение сальмонелл в клетки является примером микробзависимого фагоцитоза.


В отличие от шигелл, которые для размножения должны быстро выйти в клеточную цитоплазму, сальмонеллы многие часы остаются внутри фагосом. Через 6—24 ч в эпителиальных клетках большие вакуоли, заполненные бактериями, перемещаются к базальной мембране, освобождая сальмонеллы в основную пластинку (lamina propria) и подслизистый слой. Это сопровождается восстановлением поверхностных структур (микроворсинок) клеток и их стимуляцией к секреции интерлейкина-8 и других провоспалительных цитокинов. Аккумуляция нейтрофилов ведет к продукции простагландинов, которые в значительной степени отвечают за усиление секреции жидкости (диарею) мукозальными клетками. Можно сказать, что для сальмонелл интестинальные эпителиоциты, не являясь главной средой для размножения, служат скорее барьером, который им надлежит преодолеть. При выходе из эпителия сальмонеллы захватываются макрофагами. Испытывая микробное давление (в частности, действие продуктов SPI-1 на каспазу-1), они выделяют дополнительные порции провоспалительных цитокинов, подвергаясь апоптозу. Пребывание в фагоцитах требует от сальмонелл участия более 40 новых генов (хромосомных и плазмидных); не меньшее число генов подлежит отключению. В конечном счете верх берут защитные силы хозяина, который оказывается сильнее бактерий. Инвазия «местных» (энтеритпродуцирующих) сальмонелл практически заканчивается на этом этапе. Лишь изредка некоторые штаммы (возможно, биовары) проникают в сосудистое русло, вызывая генерализованные процессы с развитием септикопиемических состояний.
Впрочем, есть наблюдения, позволяющие усомниться в значимости интестинальной инвазии для «местных» сальмонелл. Выявлено, в частности, что для индукции энтерита необходимы лишь эффекторные белки внеклеточных бактерий. Кроме того, установлено, что различные серотипы, которые равноценно инвазируют интестинальные эпителиоциты, отличаются по энтеропатогенности. Наконец, не исключено, что некоторые сальмонеллы все-таки продуцируют энтеротоксины, меняющие кишечный транспорт электролитов и воды. Дополнительные исследования должны внести ясность в эту проблему, но пока приходится принять за основу концепцию об инвазивно-воспалительном генезе сальмонеллезной патологии.
С лабораторными целями производят культуральный анализ испражнений больного, сопоставляя его результат с данными исследования подозреваемого источника заражения. Финалом анализа является определение антигенной формулы выделенного возбудителя.
Брюшной тиф/паратифы
Иначе обстоит дело с сальмонеллами (S. typhi, S. paratyphi A, B и С), вызывающими брюшной тиф и паратифы — близкие по клинике и эпидемиологии заболевания. Это антропонозы, единственным источником которых является человек — больной или (чаще) бактерионоситель, загрязняющий фекалиями окружающую среду. Наибольшее значение имеют зараженные молоко (в меньшей степени — другие продукты питания) и вода; передача контактно-бытовым путем встречается редко.
После перорального приема патогенетически значимой дозировки возбудитель доходит до тонкого кишечника и поначалу ведет себя как «местные» сальмонеллы. Он инвазирует интестинальный эпителий, переходя в основную пластинку (lamina propria) и подслизистую оболочку, где поглощается мононуклеарными фагоцитами. С этого момента его судьба принимает иной оборот: кишечная инфекция превращается в генерализованную инвазию. Сальмонеллы переносятся в регионарные (мезентериальные) лимфоузлы, после размножения попадают в грудной лимфатический проток и далее в кровяное русло (первичная бактериемия). Микробы быстро захватываются фиксированными макрофагами печени, селезенки и костного мозга, продолжая здесь размножение с образованием гранулем и вызывая гибель макрофагов (возможно, от апоптоза). Высвобождение в кровь большого количества сальмонелл означает вторичную бактериемию и совпадает с началом серьезной клиники, т.е. знаменует окончание инкубационного периода, который в среднем продолжается 14 дней. Типичными симптомами на первой неделе являются высокая температура, головная боль, бред, нарушения сердечно-сосудистой системы и другие признаки тифозного состояния. Наблюдаются розеолезная сыпь, запор, редко диарея. Чаще всего сальмонеллы обнаруживаются (но не размножаются) в крови на протяжении первых 10 дней болезни. Это период, когда они непрерывно поступают сюда из тканей, разрушаются, высвобождая эндотоксин и вызывая синдром диссеминированной внутрисосудистой гемокоагуляции — возможно, основной патогенетический механизм брюшного тифа.
Начиная со второй недели сальмонеллы попадают из печени в желчный пузырь и отсюда с током желчи снова в тонкий кишечник. Болезнь обретает иные черты, хотя по-прежнему остается тяжелой. Процент позитивных гемокультур падает, но возрастает число копрокультур, в которых присутствует возбудитель. Сталкиваясь с сенсибилизированными Т-лимфоцитами пейеровых бляшек, сальмонеллы вызывают их стимуляцию с выделением макрофагактивирующих цитокинов, которые запускают уничтожение сальмонелл в аллергических реакциях замедленного типа. Это сопровождается повреждением тканей и возникновением язв вследствие отторжения некротических масс в центре групповых лимфатических и солитарных фолликулов. Возникает диарея («гороховый суп» с кислым запахом), которая в тяжелых случаях становится геморрагической. На третьей неделе может случиться перфорация тонкого кишечника — самая распространенная причина смерти от брюшного тифа. При неосложненном течении язвы заживают, температура постепенно снижается и больной поправляется на 4—5-й неделе (рис. 2).

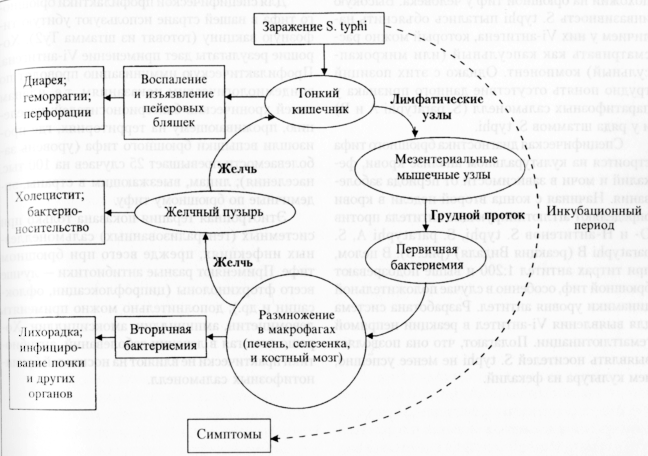
В большинстве случаев бактерии полностью элиминируются в процессе выздоровления. Однако у небольшого числа больных (2—5%) сальмонеллы остаются в желчном пузыре или (реже) в мочевыводящей системе. Половина таких людей превращаются в хронических носителей, постоянно выделяя возбудителей (чаще S. typhi) во внешнюю среду.
Безусловно, сказанное требует участия многих бактериальных генов и их продуктов. Но сегодня брюшной тиф и паратифы, бывшие когда-то весьма распространенными болезнями, встречаются редко. Это связано с расшифровкой экологии их возбудителей и разработкой методов борьбы с источниками и распространением заболевания. Большинство информации связано с наблюдениями за системой «S. typhimurium—мыши», в которой по не очень понятным причинам возникает системный процесс, похожий на брюшной тиф у человека. Высокую инвазивность S. typhi пытались объяснить наличием у них Vi-антигена, который можно рассматривать как капсульный (или микрокапсульный) компонент. Однако с этих позиций трудно понять отсутствие данного признака у паратифозных сальмонелл (S. paratyphi А и В) и у ряда штаммов S. typhi.
Специфическая диагностика брюшного тифа строится на культуральном анализе крови, фекалий и мочи в зависимости от периода заболевания. Начиная с конца второй недели в крови определяют агглютинирующие антитела против О- и Н-антигенов S. typhi, S. paratyphi A, S. paratyphi B (реакция Видаля) (рис. 3). В целом, при титрах антител 1:200 и выше подозревают брюшной тиф, особенно в случае положительной динамики уровня антител. Разработана система для выявления Vi-антител в реакции непрямой гемагглютинации. Полагают, что она позволяет выявлять носителей S. typhi не менее успешно, чем культура из фекалий.

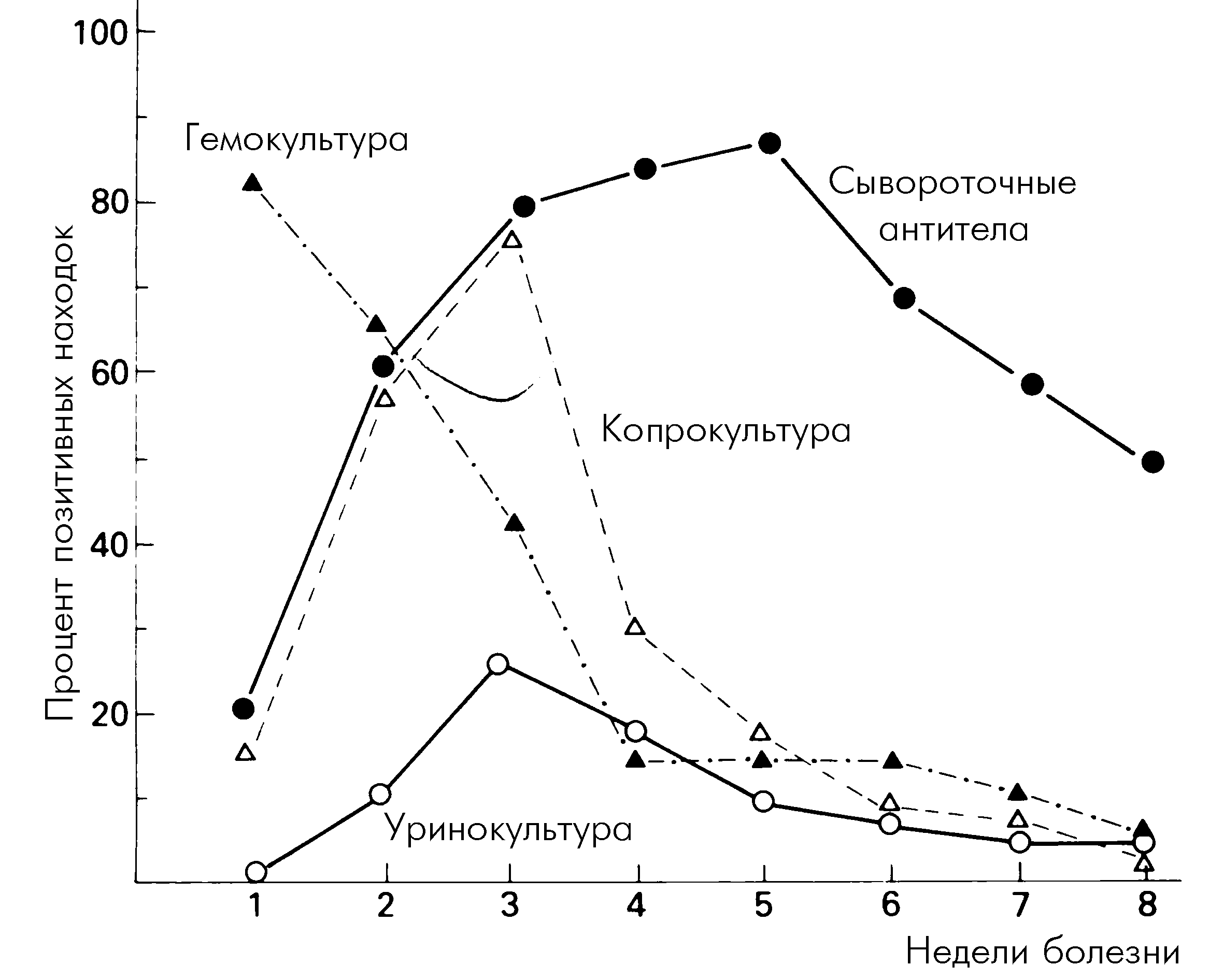
Для специфической профилактики брюшного тифа в нашей стране используют убитую тифозную вакцину (готовят из штамма Ty2). Хорошие результаты дает применение Vi-антигена. Профилактическую иммунизацию проводят по эпидемиологическим показаниям, т.е. членам семей хронических бактерионосителей; населению, проживающему на территориях, где произошли вспышки брюшного тифа (уровень заболеваемости превышает 25 случаев на 100 тыс. населения); лицам, выезжающим в страны, эндемичные по брюшному тифу.
Этиотропная терапия показана только при системных (генерализованных) сальмонеллезных инфекциях, прежде всего при брюшном тифе. Применяют разные антибиотики — лучше всего фторхинолоны (ципрофлоксацин, офлоксацин и др.); дополнительно можно применять левомицетин, ампициллин, амоксициллин. Хорошо помогая в лечении заболеваний, антибиотики практически не влияют на носителей брюшнотифозных сальмонелл.
Иерсинии
Живу не мешаю, заденут — кусаю.
- Парадоксы и логика таксономики.
- Эпидемиология.
- Экологический дуализм энтеропатогенных иерсиний.
- Серовары и патовары.
- Экологически зависимая регуляция вирулентности.
Род Yersinia включает 11 видов, но только три из них (Y. pestis, Y. enterocolitica и Y. pseudotuberculosis) имеют отношение к патологии человека. Y. pestis (возбудитель чумы) занимает высшую ступень в табеле о рангах среди возбудителей особо опасных инфекций. Полагают, что за время известных пандемий чума унесла не менее 200 млн. жизней. Первая, юстинианова, пандемия началась в Египте в 542 г. н.э. и захватила Европу, Африку и Азию. Она продолжалась до 600 г. н.э. Вторая пандемия, известная как Черная Смерть, проникла в Европу из Азии в 1330—1346 гг. Она продолжалась почти 100 лет и оказала существенное влияние на культуру, политику и систему медицинского образования. Наконец, в 1855 г. в Китае случилась третья пандемия чумы, которая постепенно распространилась на Европу и Америку. В 1894 г. в Гонконге французский бактериолог А. Иерсен выделил возбудителя чумы, дав ему название Bacterium pestis. Это открыло путь для изучения экологии агента и его распространения среди людей. Была установлена абсолютная зависимость природной чумы от циркуляции в системе «блохи—млекопитающие», прежде всего «блохи—грызуны». Человек выступает в роли случайного хозяина. Заражение происходит через укусы блох или повреждения кожи. Спустя 2—6 дней поднимается высокая температура, возникает головная боль, озноб, появляются распухшие, болезненные лимфатические узлы (бубоны), расположение которых зависит от места проникновения инфекта. Процесс носит системный характер, часто наблюдается септицемия, угрожающая жизни. Без лечения болезнь может быстро закончиться смертью от септического шока. Сегодня в развитых странах встречаются единичные случаи бубонной (незаразной) чумы среди лиц, контактирующих с животными, особенно с дикими. Они поддаются лечению антибиотиками (прежде всего стрептомицином) и редко заканчиваются вторичной (септической) пневмонией, смертельной для больного и опасной для заражения окружающих, т.е. для начала эпидемии.
Y. pseudotuberculosis и Y. enterocolitica вызывают у человека пищевые инфекции — иерсиниозы. Их природным резервуаром тоже служат мелкие грызуны, от которых они выделяются с фекалиями, заражая окружающих животных и людей. Подобно возбудителю чумы, Y. pseudotuberculosis и Y. enterocolitica обладают лимфотропностью, но это проявляется после инвазии в пейеровы бляшки терминальной части подвздошной кишки и далее в абдоминальные лимфоузлы. Процесс выглядит как энтеропатогенность. В подслизистой оболочке формируются микроабсцессы, поднимается температура, возникают абдоминальные боли, рвота и диарея с кровяными жилками. Генерализация возможна только у иммунокомпрометированных больных.
Долгое время на основании ряда общих микробиологических (мелкие грамотрицательные палочки) и эколого-патогенетических (первичные патогены млекопитающих, прежде всего, грызунов) признаков возбудителей чумы и иерсиниозов относили к пастереллам. В 1954 г. состоялось «переселение» рода Yersinia в семейство Enterobacteriaceae. С внедрением в таксономику генетических маркеров эта позиция укрепилась и сегодня выглядит незыблемой, хотя некоторые из признаков (прежде всего экологическое своеобразие) ставят иерсинии в особое положение среди энтеробактерий. Психологически трудно уравнять микроб чумы с остальными (даже наиболее патогенными) энтеробактериями, и о нем обычно говорят отдельно, пренебрегая академической систематикой. Между тем, генетическая общность Y. pestis и Y. pseudotuberculosis (в меньшей степени — Y.enterocolitica) говорит об обратном. Есть мнение, что возбудитель чумы появился около 1500 лет тому назад на основе штамма Y. pseudotuberculosis, потеряв часть генов 70-килобайтовой плазмиды и усвоив две новых плазмиды — 96 и 110-килобайтовые. C тех пор он мало изменился, сохранив исходную клональность.
Иерсинии мельче других энтеробактерий. Это овоидные палочки, близкие к коккобациллам. Типичен полиморфизм, и в мазках из культуры обнаруживаются различные формы — от кокковидных до длинных нитей. Они окрашиваются биполярно, что также нехарактерно для классических энтеробактерий. Они подвижны (кроме Y. pestis) за счет перитрихиальных жгутиков, которые образуются при температуре ниже 30°С и исчезают при 37°С. Подобно всем энтеробактериям, иерсинии не образуют спор, а капсула (гликопротеин F1, закодированный в 110-килобайтовой плазмиде) имеется только у возбудителя чумы. Факультативные анаэробы, биохимически достаточно активны, сбраживая углеводы, многоатомные спирты и пр.). Это позволяет дифференцировать их на основе классических ферментативных тестов, принятых для разграничения родовых и видовых таксонов семейства Enterobacteriaceae. В отличие от других энтеробактерий иерсинии плохо растут на лактозных дифференциально-диагностических средах, давая мелкие лактозонегативные колонии-росинки.
Важный признак — двойственность экологии Y. pseudotuberculosis и Y. enterocolitica. Инфицируя широкий круг теплокровных животных (персистенция или патогенетически значимая инфекция), они способны и к автономному (сапрофитическому) образу жизни. Эта особенность, связанная с психрофильностью, во многом определяет эпидемиологию иерсиниозов. Фенотип (в том числе продукция факторов вирулентности) энтеропатогенных иерсиний во многом зависит от температурного режима, что принято использовать для их селективного накопления в инвитровых культурах.
Источником энтеропатогенных иерсиний для человека являются синантропные грызуны (мыши, крысы), свиньи, реже крупный и мелкий рогатый скот. Заражение редко (если вообще) возникает при прямых контактах с животными или больными людьми. Как правило, инфицирование происходит из сапрофитической фазы, после размножения бактерий в обсемененных пищевых продуктах. Это связано с нарушением санитарного режима в животноводческих хозяйствах, производственных комплексах, овощехранилищах, холодильниках, столовых и пр. Размножение иерсиний в условиях холодильника на корнеплодах (бактерии попадают сюда непосредственно из почвы) и других овощах создает опасность инфицирования через свежие овощные салаты (ряд вспышек псевдотуберкулезного иерсиниоза отмечено после употребления в пищу сырой капусты, полученной из овощехранилищ). В целом пребывание энтеропатогенных иерсиний в организме хозяина можно рассматривать как факультативный (случайный) паразитизм, не обязательный для реализации жизненной стратегии возбудителя.
Подобно человеку, животные тоже страдают от иерсиний. При заражении Y. pseudotuberculosis и Y. enterocolitica у них развивается энтерит (иногда геморрагический), регионарный или системный лимфаденит, в тяжелых случаях — септицемия с поражением (гранулемы, микроабсцессы) внутренних органов (прежде всего печени, селезенки, легких). Генерализованные формы часто заканчиваются смертельно. У человека возможны те же симптомы, но к ним добавляется ряд специфических признаков, отсутствующих или не улавливаемых у животных. В целом, оба возбудителя вызывают сходные заболевания, которые предложено называть иерсиниозами. Иногда говорят о псевдотуберкулезе, или псевдотуберкулезном иерсиниозе (Y. pseudotuberculosis), и о кишечном иерсиниозе (Y. enterocolitica).
По чувствительности к антибиотикам энтеропатогенные иерсинии не отличаются от других энтеробактерий, но антибиотикотерапия абсолютно показана лишь при генерализованных формах. Местный процесс вполне обратим за счет иммунитетных ресурсов хозяина.
Первыми из иерсиний были открыты Y. pseudotuberculosis (Л. Маляссе, В. Виньяль, 1883). В 1895 г. К.Эберт, исследуя внутренние органы морских свинок, погибших при заражении этими бактериями, обнаружил узелковые образования, похожие на туберкулезные гранулемы, и ввел понятие псевдотуберкулез — термин, который закрепился в качестве видового эпитета за его возбудителем. Много лет патогенность Y. pseudotuberculosis для человека воспринималась как казуистика. Положение изменилось в 1950-х гг., когда были описаны псевдотуберкулезный вариант мезаденита и скарлатиноподобная форма иерсиниоза, впервые зарегистрированная отечественными исследователями на Дальнем Востоке (дальневосточная скарлатиноподобная лихорадка).
Псевдотуберкулезные бактерии генетически близки возбудителю чумы — около 80% гомологии ДНК. Формально этого достаточно, чтобы считать их вариантами одного вида, но гораздо меньшая болезнетворность Y. pseudotuberculosis исключает возможность подобного обобщения. Это лишь подчеркивает, как мало надо иметь бактериям для того, чтобы радикально отличаться по вирулентности от своих безвредных или менее опасных родственников. В этом отношении показательны и внутривидовые различия Y. pseudotuberculosis: величина летальной дозы для мышей у разных штаммов колеблется в миллионы раз, а из шести О-серотипов (I—VI) наиболее опасен для человека ОI.
Степень ДНК-гомологии между Y. enterocolitica и псевдотуберкулезными бактериями составляет 20—53%. Кишечный иерсиниоз (или просто иерсиниоз) — это более распространенная инфекция, чем псевдотуберкулез; последним чаще страдают животные. Болезнетворность иерсиний отчетливо ассоциирована с О-типовой принадлежностью. Сложнее всего обстоит дело с Y. enterocolitica. Она располагает не менее 30 серологическими вариантами О-антигена, причем у некоторых штаммов О-антиген неоднороден. В патологии человека главную роль играют серотипы О3; О5,27; О6,30; О8; О9. От больных животных обычно выделяются штаммы c антигенами О1; О2; О3; О5,27; О6; О7; О8; О9. Биохимически предложено дифференцировать 5 биоваров Y. enterocolitica (см. таблицу).
Биовары Y. enterocolitica
|
Признак
|
Биовары | ||||
| 1 | 2 | 3 | 4 | 5 | |
| Липаза | + | — | — | — | |
| Дезоксирибонуклеаза | — | — | — | + | |
| Образование индола | + | — | + | — | |
| Восстановление нитрата в нитрит | + | + | + | + | |
| Образование кислоты из углеводов: | |||||
| D-ксилоза | + | + | + | — | |
| сахароза | + | + | + | +/– | |
| D-трегалоза | + | + | + | — | |
Первый биовар включает две разновидности — субвары 1А и 1В, которые различаются по продукции кислоты из салицина и по пиразинамидазной активности (оба теста положительны только у субтипа 1А). К биовару 1А относятся авирулентные штаммы, выделяемые из внешней среды.
Процесс обычно ограничивается острым энтеритом/энтероколитом с диареей инвазивного типа и вовлечением в воспалительный процесс брыжеечных лимфоузлов (мезаденит, часто имитирующий клинику аппендицита — псевдоаппендицит). В этом отношении кишечный иерсиниоз не отличается от псевдотуберкулезной инфекции за исключением того, что им чаще болеют дети до 7 лет (по некоторым данным, 2/3 всех случаев иерсиниоза). Болезнь заканчивается выздоровлением через 1—2 нед, у младенцев — может продолжаться до 28 дней. Но иногда (обычно на фоне иммуносупрессии) возбудитель проникает в кровь. Иерсиниозная септицемия включает развитие абсцессов во внутренних органах (главным образом, в печени, легких и селезенке), септического артрита, менингита, эмпиемы, целлюлита и пр. Клинический полиморфизм усиливается за счет системного васкулита (полиартериит) и связанных с ним поражений кожи и слизистых оболочек. Генерализация чревата тяжелыми последствиями и часто заканчивается летально. Смертность достигает 50%.
Иерсиниозная инфекция не всегда завершается после удаления возбудителя. Возможны осложнения (лихорадка, локальные поражения), которые носят реактивный характер, т.е. развиваются при отсутствии бактерий в организме. В случае Y. enterocolitica (обычно при заражении штаммами О3 и О9) они наблюдаются у 5% больных, для Y. pseudotuberculosis — выражены у каждого третьего. Наиболее типичны поражения суставов, кожи (узелковая эритема), реже наблюдается полный синдром Рейтера (уретрит, конъюнктивит, артрит), гломерулонефрит и миокардит. Примечательно, что они ассоциированы с HLA-фенотипом: большинство больных позитивны по HLA-B27. Иммунные комплексы, изолированные от таких больных, содержат иерсиниозные антигены (артритогенные факторы?!). Обнаружены и пептиды, которые избирательно связываются с HLA-В27 и, возможно, служат мишенью для цитотоксических Т-лимфоцитов.
Еще одним предположением является гипотеза об аутоиммунной природе синдрома Рейтера и его вариантов. Дело в том, что белки наружной мембраны иерсиний располагают общими эпитопами с антигенами тканей хозяина, в частности HLA-В27, создавая возможность для образования перекрестно реагирующих антител, которые, связываясь с тканевыми антигенами, запускают воспалительный процесс. Дополнительным (или альтернативным) фактором может быть подключение аутореактивных Т-лимфоцитов, спровоцированных против HLA-В27 родственными иерсиниозными антигенами. Впрочем, ни одно из объяснений не является безупречным. Не исключено, что реактивные (иммунологичские) осложнения иерсиниоза (как и ряда других инфекций) патогенетически не однородны, хотя и маскируются под общую клинику.
Патогенез энтерального иерсиниоза складывается из нескольких стадий. После пищевого заражения (единственный путь естественного инфицирования животных и человека) возбудители закрепляются в дистальных отделах подвздошной кишки и в слепой кишке. Здесь происходит внедрение в слизистую оболочку, главным образом в М-клетки, покрывающие пейеровы бляшки. Начало этому дают три адгезина иерсиний — инвазин, Ail (от англ. attachment and invasion locus; закодированы в inv- и ail-локусах хромосомы) и YadA (от англ. yersinia adhesin A; закодирован в yadA-локусе 70-килобайтовой плазмиды). Ген inv экспрессируется при 20°С, но не при 37°С и поэтому участвует лишь в пусковой стадии колонизации. Экспрессия ail-гена начинается при 37°С. Продолжая процесс, начатый инвазином, он запускает плазмидный синтез белков наружной мембраны, в том числе фибриллярный протеин YadA. Полагают, что это главный фактор адгезии энтеропатогенных иерсиний, работающий при 37°С в организме млекопитающих. Связываясь (подобно инвазину) с b1-интегринами М-клеток, он запускает их фагоцитоз. Это объясняет склонность иерсиний к локализации в области илеоцекального угла, где сконцентрировано наибольшее количество М-клеток. Кстати Y. pestis лишены данных факторов вирулентности, что исключает заражение чумой энтеральным путем.
Главным атрибутом вирулентности иерсиний служат наружные белки, кодируемые 70-килобайтовой плазмидой, которые образуют систему секреции типа III. Большинство (около 30) из них идет на построение канала, который, пронизывая клеточную стенку бактерий, ведет к построению структуры, обеспечивающей формирование инъекционной поры в мембране эукариотической клетки. Она служит местом проникновения семи эффекторных белков, пять из которых нацелены на борьбу с фагоцитозом. Непатогенные иерсинии тоже проходят через М-клетки интестинального барьера, но, оказавшись в подслизистой оболочке, элиминируются без гистологических нарушений пейеровых бляшек. Это связано с тем, что у них не функционирует генетический аппарат, определяющий синтез белков с антифагоцитарной активностью. Такие белки получили название Yops (от англ. yersinia outer membrane proteins). Известно, что YopH, YopE, YopT, YopO/YpkA (Yersinia protein kinase A), YopJ/YopP (YopP для Y. enterocolitica) ингибируют способность фагоцитов к поглощению, подавляют их кислородзависимую бактерицидность и, в конечном счете, вызывают гибель макрофагов от апоптоза. Во многом это связано с торможением реакций в системе цитоскелета и с подавлением активности транскрипционных факторов (прежде всего NF-kB), что тормозит секрецию провоспалительных цитокинов и других медиаторов воспалительного процесса.
Как уже говорилось, 70-килобайтовой плазмидой обладают вирулентные штаммы всех трех видов иерсиний (Y. pestis, Y. psedotuberculosis и Y. enterocolitica) и все они обладают способностью к продукции Yops. Но оказалось, что этот признак тоже зависит от экологии, максимально проявляясь при 37°С в отсутствие внеклеточного Са2+ (отсюда название ряда наружных белков, контролирующих секрецию III типа — Lсr, от англ. low-calcium response). Секреция III типа — контактный феномен, который зависит от взаимодействия бактерий и эукариотических клеток и именно это является причиной реакции в условиях инфекционного процесса. Внедрившиеся в организм иерсинии проявляют агрессивность, находясь вне клеток. Этому способствует устойчивость к компонентам сыворотки, прежде всего к комплементу, которая определяется наружными белками, в частности Ail и YadA (у Y. pestis — капсульный протеин F1).
К тому, что было сказано о факторах патогенности иерсиний (Y. pseudotuberculosis и Y. enterocolitica), можно добавить информацию о дискуссии об энтеротоксине, который по механизму действия близок (если не идентичен) термостабильному энтеротоксину кишечной палочки. Сомнения относительно его патогенетических функций связаны с тем, что он почти не образуется при 37°С. Кроме того, авирулентные штаммы, лишенные плазмиды, где закодированы белки патогенности, способны продуцировать энтеротоксин, не вызывая патологии.
Много внимания уделяется иерсиниозным сидерофорам — белкам, конкурирующим за ионы железа. Допускают даже, что одним из условий, предрасполагающих к генерализации иерсиниоза, служит избыток свободного железа в организме, например при гемолитической анемии.
В целом, иерсинии следует отнести к числу наиболее многогранных патогенов. Обладая набором факторов, детерминируемых хромосомными и плазмидными генами, они являются великолепным примером бактерий, способных рационально перестраивать свой фенотип в зависимости от обстановки, добиваясь внедрения в интимные сферы гомеостаза хозяина. Это отличная модель для изучения молекулярных механизмов экологически зависимой пластичности бактерий в проекции на вполне утилитарные проблемы эпидемиологии и инфекционной патологии.
Возбудители холеры
Сто друзей — мало, один враг — много.
- Условия проявления болезнетворности холерного вибриона.
- История холерных эпидемий.
- Микробиология и серология холерного вибриона.
- Факторы вирулентности, патогенез и клиника холеры.
- Экология холерного вибриона и эпидемиология холеры.
- Лабораторная диагностика, лечение и проблемы специфической профилактики холеры.
Холерный вибрион принадлежит к роду Vibrio семейства Vibrionaceae. Сюда же относятся еще два рода, занимающих определенное место в медицинской микробиологии — Aeromonas и Plesiomonas. Все они являются бактериями с полярным жгутиком (рис. 1), живут в свежей или морской воде, в кишечнике животных и человека. Из более чем 30 видов рода Vibrio особое значение имеет Vibrio cholerae.

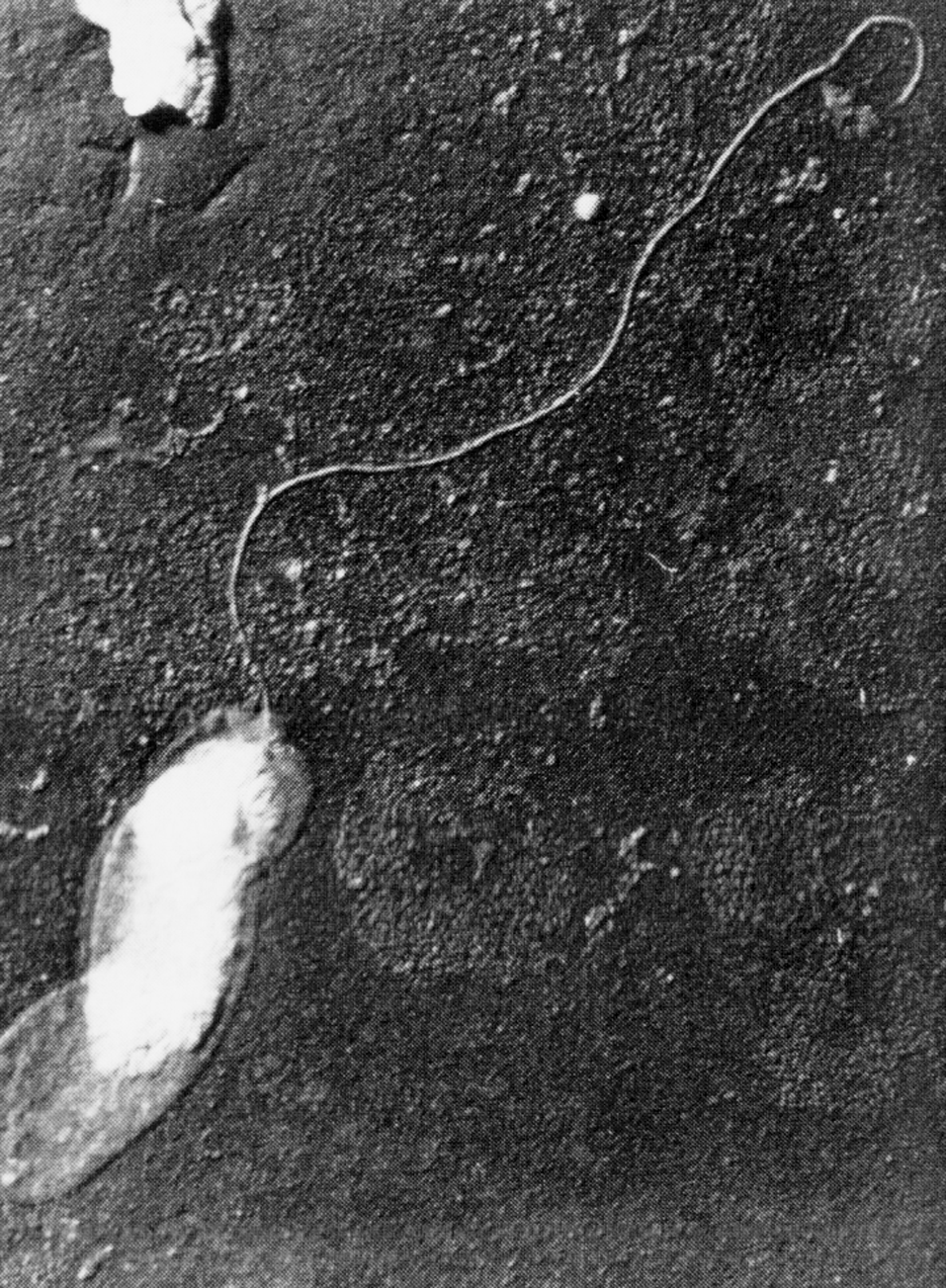
Холера — острая кишечная инфекция, вызываемая энтеротоксинпродуцирующими штаммами холерного вибриона. Она протекает в разных вариантах — от бессимптомного инфицирования до тяжелого процесса с внезапной профузной диареей, ведущей к гиповолемическому шоку, ацидозу и смерти. Это антропоноз, поражающий только человека; мишенью для возбудителя служат энтероциты тонкого кишечника. Здесь холерные вибрионы находят оптимальные условия для быстрого размножения (щелочная среда, продукты переваривания белков) и даже вытесняют другие микробы.
Вибрион поступает в кишечник через кислую среду желудка, которая служит для него мощным барьером. Являясь ацидочувствительным, он преодолевает эту преграду только при массированном заражении (106 токсигенных вибрионов не вызывают симптомов у взрослых добровольцев, если они принимаются без пищи), хотя реальная заражающая дозировка может быть ниже. Вибрионы ускользают от действия соляной кислоты не только при нарушениях секреторной деятельности желудка (гипоацидный гастрит), но и при его нормальном функционировании. Это происходит, если холерный вибрион находится внутри пищевого комка (который защищает его от кислот) или при поступлении в желудок вместе с водой (особенно щелочной), которая быстро выбрасывается в кишечник, снижая концентрацию соляной кислоты. При приеме тех же токсигенных вибрионов в количестве 106 вместе с пищей развитие холеры наблюдали у 80—100% добровольцев. Тщательные микробиологические исследования, проведенные в Бангладеше, показывают, что клинически значимая дозировка V. cholerae составляет 102—104 клеток. Опыты на добровольцах, которые проводились на фоне подавления кислотности желудка бикарбонатом натрия, в целом подтверждают эти наблюдения. Диарейная болезнь у большинства субъектов развивалась после приема 103—104 холерных вибрионов, но заболевание протекало менее тяжело, чем от более высоких дозировок.
Холера известна с незапамятных времен. Она возникла в Южной Индии, в низовьях рек Ганга и Брахмапутры, т.е. на территории, сегодня известной как государство Бангладеш. До начала ХIX столетия холера оставалась эндемичной для данного региона, и только в 1817 г. с развитием миграции населения и расширением людских контактов произошло распространение инфекции на соседние регионы и отдаленные страны, включая Европу и Америку. Холера стала пандемически опасной болезнью, борьба с которой требует огромных средств. Ежегодные вспышки уносят жизнь около 120000 человек, но еще больше людей страдает от тяжелых форм холеры. Строгое соблюдение гигиенических мер безопасности, основанных на знании экологии возбудителя, ограничивает распространение холеры регионами Индостана, Южной Азии, Африки и Латинской Америки. Однако с опасностью заноса холеры приходится считаться всем странам, включая Россию. Не случайно холеру называют в числе «вновь появляющихся» инфекций (emerging and reemerging infections), угрожающих многим развивающимся странам.
С 1817 г. человечество знало восемь пандемий холеры, унесших (и продолжающих уносить) огромное число человеческих жизней. Во время пятой из них, в 1883—1884 гг., Р. Кох изолировал ее возбудителя, V. cholerae, который сегодня отнесен к биотипу «классический». В 1961—1962 гг. началась новая, седьмая по счету пандемия холеры, вызванная другим биотипом V. cholerae О1 — Эль-Тор. Он вырвался за пределы своего эндемичного очага на острове Сулавеси в Индонезии и обычным путем, через государства Юго-Восточной Азии, охватил Средний Восток, африканские страны, южные районы бывшего СССР и даже достиг Европы (Португалия, Испания, Франция), Америки и Японии. Пандемия прекратилась к 1993 г., оставив после себя новые очаги эндемичной холеры в Азии и Африке. В 1962 г. ВОЗ признал инфекцию, обусловленную вибрионом Эль-Тор, холерой, включив ее в группу карантинных заболеваний. Это произошло всего через четыре года после заявления о том, что холера исчезает, а инфекцию Эль-Тор нельзя считать холерой.
В 1992—1993 гг. в Индии и Бангладеше зарегистрирована эпидемия «новой» холеры, вызванной штаммом О139 V. cholerae. С тех пор вспышки холеры О139 появлялись в Пакистане, Афганистане, Непале, Китае, Таиланде и других странах, соседствующих с первоначальным очагом. На этот раз ВОЗ поступила осторожнее, быстро признав в V. cholerae О139 возбудителя холеры с пандемическим характером распространения.
Итак, возбудителями современной холеры являются представители серогруппы О1 (V. cholerae, биотип classiс и V. cholerae, биотип El-Tor, почти вытеснивший первого из них) и О139 (V. cholerae O139, син. Bengal). Вариант Эль-Тор отличается от классического биотипа по своей гемолитической активности (гемолиз бараньих эритроцитов) и резистентности к полимиксину В. Их можно дифференцировать также по чувствительности к специфическим фагам.
О1-групповые штаммы (классический и Эль-Тор) имеют антигенные детерминанты А, В и С, которые делают возможным их дальнейшее типирование. Сочетание АВ-субъединиц называется субтипом Огава, АС — серотипом Инаба, АВС (встречается редко) — серотипом Хикошима (рис. 2). Эти особенности используются в эпидемиологических исследованиях для определения сходства или различия штаммов, спровоцировавших холеру. V. cholerae О139 очень похож на серовар О1, что допускает его происхождение от биотопа Эль-Тор. Подобно другим не-О1 холерным штаммам, он образует капсулу, которая скорее всего не участвует в болезнетворном процессе.


V. cholerae является короткой, слегка искривленной, высоко подвижной грамотрицательной палочкой. У него один полярно расположенный Н-жгутик, который серологически идентичен у всех холерных вибрионов. По липополисахаридному О-антигену холерные вибрионы разделяют более чем на 150 серотипов (серогрупп), которые можно отличить не только по этому признаку, но и по генотипическим деталям, отражающим степень вирулентности. По мере обнаружения новых О-серотипов возникло противоречие, связанное с признанием того, что холеру вызывают представители лишь одного О-серотипа V. cholerae — О1. Пришлось принять компромиссное решение — оставив неизменным название вида (V. cholerae), остальные штаммы именовать не-О1-серотипами, или неагглютинирующимися. Оказалось, что не-О1-штаммы способны вызывать диарейные (даже холероподобные) инфекции у человека (спорадические и небольшие вспышки) за счет плохо понятных факторов, не связанных с продукцией холерного энтеротоксина. Более того, с 1992 г. началось пандемическое распространение холеры, связанной с энтеротоксигенным штаммом О139 V. cholerae, который, выйдя из южной части Индийского полуострова, обрел черты эпидемичности.
V. cholerae не образует спор. Поэтому он весьма чувствителен к физическим и химическим агентам. Погибает при умеренно высоких температурах (10 мин при 55оС) и столь же легко уничтожается обычными дезинфектантами. Факультативный анаэроб со склонностью к аэробным условиям существования. Восстанавливает нитраты в нитрит, и это служит стимулом для анаэробного роста при наличии ферментируемого субстрата, например глюкозы или сахарозы. Вообще, подобно другим вибрионам, V. cholerae отличается неприхотливостью и может быть выращен на обычной пептонной воде. Негалофилен, или галотолерантен, что означает его способность выживать и развиваться на средах, не содержащих или включающих до 3% NaCl. Оксидазоположителен, что отличает V. cholerae от энтеробактерий, которые тоже выделяются из кишечника человека и животных. Оптимальная температура роста — 30—37оС, но размножение может быть получено в диапазоне от 16 до 42оС. Это служит лимитирующим фактором для накопления V. cholerae в окружающей среде (прежде всего в воде), способствуя сезонности холерных эпидемий. Продуцирует комплекс экстрацеллюлярных ферментов, которые расщепляют ДНК, желатин, хитин, белок, лецитин, утилизируют крахмал и многие углеводы (сбраживание до кислоты). Это сближает V. cholerae с другими вибрионами, родами Aeromonas и Plesiomonas (которые тоже способны вызывать заболевания у человека) и больше годится для взаимоотношений с внешней средой, чем с человеком.
В патогенезе инфекции серьезную роль играет повышенная чувствительность V. cholerae к кислотам и несколько необычная резистентность к щелочам и солям желчных кислот. Во-первых, это требует от холерного вибриона высокой дозировки для того, чтобы преодолеть кислую среду нормального желудка и инициировать инфекцию. Во-вторых, способствует выживанию бактерий, оказавшихся в условиях повышенного содержания солей желчных кислот и щелочной среды слизистой тонкого кишечника. Относительная толерантность холерных вибрионов к высоким рН (до 9,6) и таким ингибиторам бактериального роста, как тиосульфат, соли желчных кислот и теллурит, широко используется в приготовлении селективных питательных сред для культивирования холерного вибриона. Холерные вибрионы составляют первую группу (из шести) по Хейбергу, т.е. разлагают маннозу и сахарозу, но инертны к арабинозе. В предварительной идентификации V. cholerae ключевым шагом служит позитивный оксидазный тест: он отличает их от энтеробактерий, загрязняющих водоемы.
Патогенез холеры — сложный процесс, который зависит от комплекса факторов, обеспечивающих колонизацию эпителия (энтероцитов) тонкого кишечника и продукцию энтеротоксина, разрывающего ионный транспорт в интестинальных клетках. Хотя образование холерного токсина, закодированного в ctxAB-генах (англ. cholera toxin, A и В субъединицы токсина), напрямую связано с явлениями диареи, развитие холеры возникает вследствие синергического эффекта многих генов, включая гены колонизационных факторов, прежде всего tcp (англ. toxin coregulated pili). Кстати, холерный энтеротоксин был открыт лишь в 1959 г. на основе теста с изолированной кишкой кролика в независимых опытах De и Dutta (рис. 3). До этого считалось, что главным в патогенезе холеры является эндотоксин, всасывающийся в кровь из кишечника.

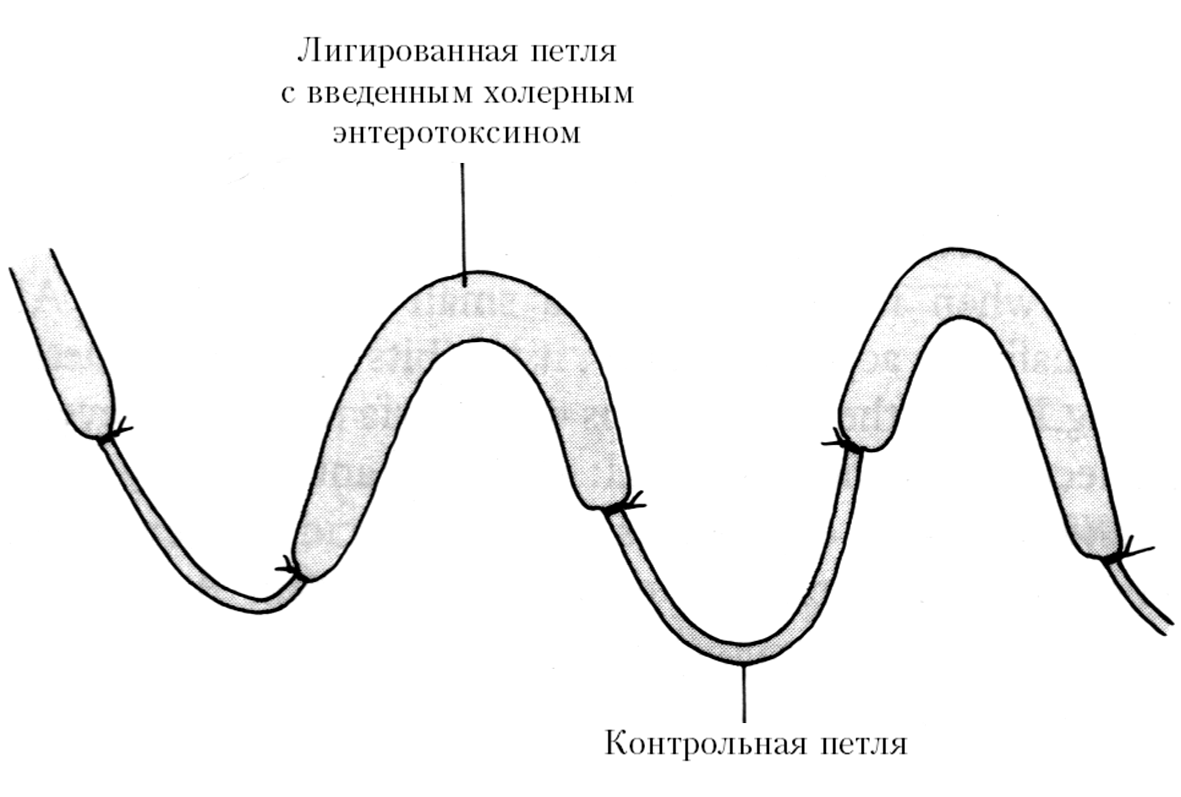
Основные гены вирулентности находятся в трех хромосомных кластерах (островах патогенности), два из которых привносятся в клетку при помощи умеренных нитевидных бактериофагов. Сегодня известно, что координированная экспрессия генов ctxAB и tcp контролируется в составе регулона тремя регуляторными генами toxS, toxT и toxR, среди которых последний является глобальным геном-регулятором. Отсутствие или повреждение гена toxR у штаммов, имеющих структурные гены ctxAB и tcp, обусловливает авирулентность или слабую вирулентность V. cholerae, резко (более чем в 100 раз) снижая продукцию холерного энтеротоксина и токсинкорегулируемых пилей. Присутствие в геноме вибрионов указанных генов и их эффективная экспрессия обеспечивают развитие инфекционного процесса при заражении человека.
Ген tcp (точнее его главный компонент tcpА) рассматривается как инициирующий элемент в каскаде генов tcp-острова патогенности. Его экспрессия определяет синтез TCP-пилей, которые являются ведущим фактором микробной колонизации человека. Кроме того, ТСР-пили служат рецептором для фага, вносящего гены холерного токсина (CTX), создавая условия для горизонтального приобретения этого важнейшего атрибута холерного вибриона. Полагают, что такой обмен может происходить внутри тонкого кишечника, где создаются оптимальные условия для взаимодействия бактерий с лизогенными фагами. Важно, что многие клинические изоляты V. cholerae O1 и О139 несут множество копий СТХ и, следовательно, ctxAB-оперонов. Это повышает вирулентность штаммов, делая их более агрессивными.
Молекула холерного токсина состоит из белковых субъединиц, которые собираются вместе в периплазматическом пространстве — одной А и пяти В. Компонент В служит рецептором токсина, связываясь с GM1-гликозидами на поверхности эпителиальных клеток тонкого кишечника. Оказавшись внутри энтероцитов, холерный токсин высвобождает свою субъединицу А (точнее А1) и находит молекулярную мишень (белок Gs), действие на которую ведет к холерной диарее. Этот эффект происходит в результате АДФ-рибозилирования Gs-белка, как это наблюдается при действии дифтерийного токсина на фактор элонгации EF-2 (см. «Возбудитель дифтерии»). Такой механизм лишает Gs-белок гуанинтрифосфатазной активности, снимая контроль с торможения аденилатциклазы. Создается повод для бесконтрольного синтеза циклического аденозинмонофосфата (цАМФ) и через это — к извращениям в обмене электролитами и потере воды.
Холера (особенно холера Эль-Тор) часто протекает в бессимптомной или легкой форме. Это создает круг носителей вибриона, которые во время эпидемий сбрасывают его в окружающую среду (в водные резервуары, на пищевые продукты), расширяя потенциальные источники инфекции. В очагах эндемичной холеры V. cholerae, подобно другим вибрионам, постоянно находится в качестве нормального резидента водных водоемов, и его присутствие не обязательно связано с фекальным загрязнением. Природные условия не только благоприятствуют здесь выживанию холерного вибриона, но и содействуют горизонтальному обмену генетическими элементами, например приобретению лизогенного фага СТХf, превращающего бактерии в формы, продуцирующие холерный энтеротоксин. Это может придавать эндемичной холере эпидемичные формы, поддерживая сезонный характер заболеваемости. Впрочем, к аналогичному результату способны вести и взаимоотношения холерного вибриона с водным зоо- и фитопланктоном. Обсуждается вопрос о персистенции бактерий в живой, но не культивируемой форме, хотя возможность реверсии в обычную форму для холерного вибриона считается маловероятной.
Холера часто ведет к развитию водной диареи с дегидратацией (обезвоживанием) организма и потерей электролитов. Активация холерным токсином аденилатциклазы ведет через 1—5 дней к усиленной секреции энтероцитами воды и электролитов в просвет тонкой кишки. Это обусловливает появление характерной водянистой диареи (вид рисового отвара без фекального запаха) и рвоты, объем которых может достигать до 30 л и более в сутки. Показательно, что заболевание не сопровождается воспалительной реакцией, т.е. действие холерного токсина ограничивается фармакологическим гормоноподобным эффектом. Возбудитель размножается в поверхностных слоях эпителия, прикрепляясь с поверхности энтероцитов Tcp-пилями и продуцируя большое количество экзотоксических субстанций. Определенное значение играют подвижность вибрионов (неподвижные мутанты менее вирулентны, чем дикие бактерии), муциназа, нейраминидаза и другие ферменты и токсины, которые есть у всех патогенных вибрионов, но все они не идут в сравнение с патогенетическим значением Tcp-пилей и холерного энтеротоксина.
Выделяют эндемичную и эпидемичную (пандемичную) холеру. Первая из них ограничена зоной первичного очага инфекции, прежде всего южной частью Индийского полуострова (низовья Ганга и Брахмапутры). Эндемичная холера предполагает постоянную заболеваемость (больше в летне-осенний период), прерываемую всплесками холерной инфекции. По существу, речь идет о местной эпидемии, т.е. о том, что можно назвать «эндемичной эпидемией холеры». Персистируя в водных экосистемах независимо от человека, холерный вибрион вызывает первые случаи болезни при заражении из водных резервуаров. Появление больных приводит к инфицированию водоемов вирулентными холерными вибрионами, обусловливая взрывной подъем заболеваемости. Таким образом, вода поверхностных водоемов имеет многогранное значение в качестве резервуара и фактора передачи возбудителя инфекции. Это основной путь мобилизации холерных вибрионов, приводящий к эпидемическому распространению болезни, дополняемый алиментарной и контактно-бытовой передачей инфекта. Холера является заболеванием густо населенных районов с низкими гигиеническими стандартами. Инфекция упорно присутствует в коллективах, снабжаемых коммунальной водой, использующих каналы и реки для купания и хозяйственных нужд. Поэтому холера остается эндемичной инфекцией для Бангладеша и многих частей Азии и Африки.
Возникает вопрос о происхождении так называемых эпидемичных штаммов холерного вибриона. Речь идет о токсигенных клонах, которые обусловливают вспышки заболеваний вне эндемичных очагов инфекции. Их природа остается неясной, хотя на этот счет есть предположения. Можно, например, думать о неоднородности (а, следовательно, о возможности отбора) клонов с более высокой устойчивостью в окружающей среде, более выраженной способностью к выделению из организма, повышенной токсигенностью и условиями для ее экспрессии. Генетическая неоднородность штаммов V. cholerae неоднократно подчеркивалась различными исследователями, и, по-видимому, правильнее говорить о клонах с неодинаковой вирулентностью, чем о вариантах с разным эпидемическим потенциалом. Повышение вирулентности штамма будет способствовать его эпидемической (пандемической) значимости. Что касается механизмов появления высоковирулентных клонов, то здесь могут участвовать подвижные гены сtxAB, tcpA, toxR, hap и другие, работающие в масштабе регулона и обеспечивающие одновременную экспрессию различных признаков вирулентности V. cholerae.
Лабораторная диагностика холеры служит клиническим и эпидемиологическим целям. Главным является культуральный метод, т.е. выделение и идентификация чистой культуры возбудителя. Для этого используют посев исследуемого материала (фекалии, рвотные массы, вода, пищевые продукты) на элективные среды с последующим анализом колоний на морфологические и ферментативные признаки, характерные для холерного вибриона. В специализированных лабораториях изучают принадлежность вибриона к О-серогруппам. Для серогруппы О1 исследуют отношение к биотипу (классический, Эль-Тор) и серотипу (Огава, Инаба, Хикошима). При необходимости определяют чувствительность к антибиотикам.
Принцип лечения холерных больных сводится к немедленному восполнению дефицита воды и электролитов с помощью глюкосолевых растворов — регидратация и реминерализация. При легких и среднетяжелых формах бывает достаточной оральная терапия; тяжелобольным назначают внутривенные вливания сходных растворов. Оральный изотоничный раствор включает глюкозу, хлорид натрия, хлорид калия и бикарбонат натрия (см. «Ротавирусы»). Глюкоза служит проводником для всасывания воды и электролитов, применение одной воды не дает позитивных результатов. Использование регидратации резко снизило число смертельных исходов холеры — до 1—5%.
Если оральная и внутривенная регидратация не прекращают диареи, больным назначают антибиотики (тетрациклины, хлораминфеникол, фторхинолоны), которые, действуя на холерный вибрион, способствуют остановке процесса.
Опыты на добровольцах подтверждают эпидемиологические наблюдения о том, что заболевание холерой создает иммунитет, который сохраняется по крайней мере три года. Это оправдывает попытки разработать вакцину против холеры. Парентеральные препараты оказались неэффективны, поэтому современное внимание направлено на получение оральных вакцин. Испытание живых аттенуированных препаратов в комплексе с компонентом В холерного токсина или без него позволили добиться определенного результата. Однако вакцина дает умеренные осложнения (диарея) и мало эффективна в детском возрасте. Последнее обстоятельство особенно печально, так как младенцы и дети служат группой повышенного риска, особенно в зонах эндемичной холеры. Поэтому вопрос о противохолерной вакцине остается делом будущего. Главное в профилактике холеры — неспецифические мероприятия, направленные на предупреждение контакта с возбудителем.
Хеликобактер пилори
Ценность гипотезы определяется тем, насколько активно
она побуждает к тому, чтобы ее опровергнуть.
Ф.М. Бернэт
- Новое в таксономике.
- Парадоксы экологии.
- Факторы и условия выживания в кислой среде желудка.
- Первичный или оппортунистический патоген?
- Механизмы колонизации.
- Гастродуоденальная патология: пусковые и поддерживающие начала.
- Диагностические критерии.
- Терапия и патогенетический приоритет H. pylori.
В 1983 г. австралийскими микробиологами B. Marshall и R. Warren было сделано неожиданное открытие. По сути в забытой культуре из эндоскопического биоптата пилорического отдела желудка были обнаружены неизвестные бактерии, давшие начало новому родовому таксону Helicobacter и его типовому виду H. pylori (лат. helix — спираль, pyloris — привратник). Этой случайной находке было суждено изменить представления о природе одного из самых распространенных и опасных заболеваний пищеварительного тракта человека — язвенной болезни желудка и двенадцатиперстной кишки. Сообщения о присутствии спиралевидных (спирохетоподобных) и изогнутых бактерий на слизистой оболочке желудка человека и животных неоднократно публиковались и ранее, но именно Marshall и Warren впервые выделили их в чистой культуре и постулировали причинную связь с гастродуоденальной патологией. В 2005 г. за свое открытие Marshall и Warren были удостоены Нобелевской премии.
H. рylori представляет собой грамотрицательную тонкую извитую (в форме запятой или S-образную) палочку длиной 1—5 мкм. На одном из ее концов имеется несколько жгутиков, обеспечивающих подвижность. Споры и капсулу не образует, растет при низкой концентрации кислорода (микроаэрофил). Относится к неферментирующим бактериям, получая энергию за счет окисления аминокислот и производных цикла трикарбоновых кислот; углеводы не утилизирует. Растет на обычных кровяных средах при повышенной влажности, но для получения колоний требуется довольно много времени — 3—5 сут, иногда более. В сочетании с микроаэрофильностью это затрудняет выделение H. pylori, вынуждая пользоваться методами, основанными на определении макромолекулярных продуктов бактерий (антигены, нуклеиновые кислоты) и серодиагностике. Важным с экологической (и, возможно, патогенетической) точки зрения свойством является уреазная активность, т.е. способность к расщеплению мочевины. Это вызывает защелачивание вокруг бактерий (результат образования аммиака и двуокиси углерода), обеспечивая выживание в условиях кислой среды желудка. Кроме высокоактивной уреазы H. pylori обладает оксидазой и каталазой. Они необходимы для получения энергии (оксидаза) и защиты от реактогенных метаболитов кислорода (каталаза), которые продуцируются нейтрофилами, инфильтрирующими зону хеликобактерной колонизации.
Открытие H. pylori способствовало обнаружению группы родственных бактерий, и сегодня род Helicobacter включает около десятка видов, изолированных из желудка и нижних отделов пищеварительного канала различных животных. Отличаясь в деталях, они генетически близки между собой и хорошо адаптированы к необычным условиям, с которыми им приходится сталкиваться в желудочном содержимом. Это говорит о том, что разные виды Helicobacter имеют общее происхождение и эволюционировали вместе со своими хозяевами в ранге нормальной или почти нормальной микрофлоры. Для вида H. pylori человек — единственный естественный хозяин, а его желудок — уникальная экологическая ниша, в которой H. pylori способен длительно (возможно, пожизненно) персистировать, не вступая в патогенетически значимый конфликт с хозяином.
В желудок попадает множество бактерий, но они быстро погибают, не выдерживая кислой среды, либо проходят транзитом в кишечник. Лишь при гипоацидозе гастральная слизистая оболочка заселяется интестинальной (т.е. типичной для кишечника) микрофлорой, но это уже вторичная инфекция, результат патологии. H. pylori — единственный представитель специфической микрофлоры желудка, который способен колонизировать его нормальную слизистую оболочку и надолго задерживаться там. Из всех отделов желудка H. pylori отдает предпочтение антральной части привратника. Именно здесь он наиболее интенсивно колонизирует эпителий, внедряясь в слой покрывающей его слизи (рис. 1).


Это, очевидно, связано с тем, что в привратнике (в отличие от тела и дна желудка) мало клеток, секретирующих соляную кислоту, что облегчает выживание бактерий на поверхности эпителия, защищенной от кислого содержимого желудка толстым слоем слизи. Большинство бактерий свободно плавает в ней, и лишь около 20% прикреплены к эпителиоцитам. Возможно, это и есть та естественная экологическая ниша, которая эволюционно закреплена за H. pylori.
H. рylori — один из самых распространенных инфекционных агентов человека: он присутствует в желудке около половины населения Земного шара. Находясь здесь, H. pylori не приносит (или почти не приносит) вреда хозяину. С этой точки зрения его следует отнести к бактериям-оппортунистам, условная патогенность которых сдерживается хозяином. Но иногда ситуация выходит из-под контроля: вне пределов своего естественного биотопа (антрального отдела желудка) H. pylori вызывает деструктивный процесс, поддерживаемый другими факторами. Ими могут быть повреждения слизистой оболочки соляной кислотой или экзогенными ирритантами, гастральная метаплазия эпителия 12-перстной кишки и прочие, до сих пор не ясные причины. Кроме того, не все штаммы H. pylori обладают одинаковой вирулентностью (см. ниже).
Первое указание на то, что H. pylori имеет отношение к гастродуоденальной патологии было встречено скептически, с активным нежеланием отойти от традиционной (хотя и противоречивой) позиции, согласно которой пептические язвы обусловлены избытком соляной кислоты в желудочном соке. Но сомнения вскоре рассеялись, и сегодня мало кто отрицает хеликобактерную природу большинства случаев язвенного поражения гастродуоденального тракта. Вместе с тем очевидно, что H. pylori действует не в одиночку, а в сочетании с другими, неизвестными или малоизученными факторами. Об этом говорит то, что большинство cлучаев хеликобактерной инфекции не сопровождается язвенными симптомами, а примерно 30% пептических язв желудка не связаны с присутствием H. pylori, по крайней мере в зоне эрозии (это особенно типично для антрального отдела). Неоднозначно и отношение к ассоциированному с хеликобактером воспалению антральной зоны желудка — так называемому хроническому активному гастриту. Он называется так потому, что, наряду с мононуклеарными клетками (моноцитами, лимфоцитами, плазматическими клетками), в инфильтрате присутствуют нейтрофилы, т.е. элементы хронического воспаления сочетаются здесь с признаками острого процесса. В целом это скорее гистологическое понятие, так как клинические симптомы обычно отсутствуют, а атрофия эпителиального покрова слизистой оболочки (атрофический гастрит), если и развивается, то очень медленно. Впрочем, для дуоденальных язв эти противоречия сглажены: они более надежно ассоциированы с хеликобактерным гастродуоденитом. Многие склоняются и к тому, что длительная (30—40-летняя) воспалительная реакция, ведущая к метаплазии и атрофии эпителия слизистой оболочки, существенно (по некоторым данным в 6 раз) повышает риск развития рака желудка.
Обобщенная формула гласит: хеликобактерная инфекция — необходимое, но недостаточное условие для развития язвенной болезни. Основные доказательства в пользу этиологической значимости H. pylori сводятся к следующему: 1) за исключением случаев, когда язвы возникли после поступления в желудок раздражающих веществ (например, нестероидных противовоспалительных препаратов), более 95% больных с дуоденальными язвами и более 80% больных с язвами желудка инфицированы H. pylori, что существенно выше уровня инфицированности здоровых людей; 2) у лиц, зараженных H. рylori, язвы 12-перстной кишки развиваются гораздо чаще, чем у неинфицированных людей; 3) по сравнению с традиционной антисекреторной терапией элиминация H. pylori при помощи антибактериальных средств гораздо надежнее предупреждает рецидивы язвенной болезни.
Уникальная способность H. pylori колонизировать слизистую оболочку желудка определяется как минимум тремя свойствами, сочетание которых встречается только у бактерий данного вида:
- нейтрализация кислой среды желудка;
- активное внедрение в слой слизи, покрывающей эпителий;
- избирательное закрепление на эпителиоцитах слизистой оболочки.
Как уже говорилось, ослабление кислой реакции достигается благодаря ферменту уреазе, по активности которой хеликобактеры не имеют себе равных среди бактерий. В присутствии мочевины H. pylori сохраняет жизнеспособность в течение 90 мин при рН=2,0. Этого вполне достаточно, чтобы успеть внедриться в слизь, защищающую от соляной кислоты. Проникновение в вязкий слой слизи достигается благодаря штопорообразной подвижности, обусловленной спиралевидной формой и полярными жгутиками. Неподвижные варианты H. pylori обладают слабой колонизирующей активностью и в отличие от подвижных бактерий не вызывают хронического гастрита.
С уреазой H. pylori связан еще один сюрприз: значительная часть фермента находится на поверхности бактерий и высвобождается путем генетически запрограммированного лизиса бактериальных клеток. Такого рода альтруистический аутолиз, впервые обнаруженный у H. pylori, повышает жизнеспособность соседних бактерий в неблагоприятных условиях.
H. pylori прикрепляется к клеткам многих тканей. Однако в условиях естественной инфекции он отдает явное предпочтение эпителию гастрального типа. Так, в 12-перстной кишке бактерии заселяют зоны гастральной метаплазии; прилежащие к ним клетки интестинального типа не колонизируются. Соответственно, в желудке H. pylori избегает участков слизистой оболочки с интестинальной метаплазией, т.е. с эпителием кишечного типа. Бактериальные факторы, определяющие адгезию, точно не установлены, хотя имеется несколько кандидатов среди фибриллярных (пилеподобных) структур с гемагглютинирующей активностью. Они связываются с мембранными гликолипидами, протеогликанами, сиалированными ламинином (входящим в состав межклеточных контактов и базальной мембраны) компонентами слизи. Эффект специализированных адгезинов усиливается липополисахаридом наружной мембраны. Впрочем, по аналогии с некоторыми видами хеликобактеров животного происхождения закрепление на эпителиоцитах может быть не обязательным условием для колонизации слизистой оболочки. Бактерии способны персистировать в слое слизи, покрывающей эпителий (первичная адгезия). Инвазия нехарактерна, и лишь единичные бактерии подвергаются эндоцитозу, сохраняясь внутри эпителиоцитов.
Специфика адгезинов не исчерпывает комплекса факторов, определяющих высокую избирательность H. pylori в отношении человека. Подобно другим бактериям, важнейшим фактором роста для хеликобактеров является железо. Они добывают его непосредственно из лактоферрина мукоидного секрета, используя для этого лактоферринсвязывающий белок наружной мембраны. Лактоферриновые рецепторы H. pylori обладают удивительным свойством: фиксируя человеческий лактоферрин, они не взаимодействуют с аналогичным белком животных. Это лишний раз подчеркивает, что базисные условия для своего развития H. pylori получает только в организме человека.
Обосновавшись на слизистой оболочке, H. pylori начинает продуцировать факторы, которые запускают и поддерживают воспалительную реакцию в стенке желудка. Имеется прямая корреляция между количеством бактерий на слизистой оболочке и интенсивностью воспалительной реакции. Сама зона колонизации (т.е. поверхность инфицированной слизистой оболочки) остается вне сферы действия воспалительных клеток, которые локализуются в субэпителиальном слое слизистой оболочки. Это позволяет бактериям ускользать от прямой атаки, способствуя их персистенции. H. pylori обладает комплексом факторов, активирующих нейтрофилы, эпителиальные клетки, моноциты и Т-лимфоциты. Кроме того, в очаге воспаления выделяются флогогенные медиаторы (интерлейкины-1, -6, -8, туморонекротический фактор, гамма-интерферон, фактор активации тромбоцитов и пр.), которые поддерживают воспалительный процесс.
Деструкция слизистой оболочки с развитием язвенных поражений происходит сравнительно редко, и ее пусковые механизмы гипотетичны. У H.pylori обнаружены токсины, повреждающие эпителиоциты in vitro. Но они продуцируются не всеми штаммами, выделяемыми от больных с язвенной патологией, и их патогенетическое значение требует дополнительных обоснований. Это, в частности, справедливо для вакуолизирующего (VacA) и CagA (англ. cytotoxin-associated antigen) токсинов. Их продукция колеблется между штаммами и не может отвечать за всю вирулентность. То же самое справедливо для факторов с антипролиферативной активностью, которые, возможно, ослабляют репарацию поврежденного эпителия. Приводятся доводы в пользу цитотоксичности фосфолипазы и продуктов уреазной активности (аммония). Установлено деполимеризующее действие ферментов H. pylori на слизь, покрывающую эпителий желудка. Истончение слоя слизи делает эпителиоциты более уязвимыми для соляной кислоты.
В целом, главной остается классическая трактовка, согласно которой возникновение пептических язв связано с действием соляной кислоты на фоне снижения регенеративных потенций эпителия и факторов, нейтрализующих кислотное содержимое желудка. Вопрос лишь в том, каким образом H. pylori помогает индукции и реализации данного механизма. Можно, в частности, думать о суммарном воздействии микробных и эндогенных факторов, тем более, что в биоптатах из зоны активной язвы 12-перстной кишки H. pylori обнаруживается лишь у 15,9—56% больных. Это ставит под сомнение приоритет микробных продуктов в прямом повреждении эпителия, побуждая к более сложным патогенетическим схемам. Допускается, например, что H. pylori прямо или косвенно (через цитокины и другие медиаторы воспаления) влияет на физиологическое равновесие в системе клеток слизистой оболочки желудка, секретирующих соматотропин, гастрин и соляную кислоту. Возникающий дисбаланс в секреции соляной кислоты в сочетании с предрасполагающими (в том числе микробными) факторами ведет к повреждению слизистой оболочки.
Среди возможных причин тканевой деструкции рассматриваются и нейтрофилы. H. pylori побуждают к продукции хемоаттрактантов поли- и мононуклеарных фагоцитов, которые (прежде всего интерлейкин-8) выделяются мукозальными эпителиоцитами при контакте с живыми H. pylori, особенно с cag-позитивными вариантами. Из водных экстрактов бактериальных клеток и продуктов метаболизма выделен белок, который усиливает интегринзависимое взаимодействие нейтрофилов с эндотелиоцитами и стимулирует их кислородзависимый метаболизм. Признаки активации нейтрофилов отмечаются непостоянно и не у всех больных (это может быть связано с особенностями хеликобактерных штаммов), но в ряде случаев эпителий подвергается агрессии со стороны оксидантов, катионных белков и протеолитических ферментов стимулированных нейтрофилов. Это создает опасность изъязвления, а при частом и длительном повторении — атрофии эпителиоцитов.
Предложена и иммунологическая концепция хеликобактерной патологии. H. pylori располагает антигенами, которые перекрестно реагируют с эпителиоцитами слизистой оболочки желудка, что допускает образование аутоантител и аутоагрессивных Т-лимфоцитов. Однако иммунопатогенетическая (аутоиммунная) трактовка деструктивного процесса не может быть принята для объяснения язвенного поражения 12-перстной кишки, так как ее эпителиоциты лишены общих эпитопов с антигенами H. pylori. Иммунопатогенетическая гипотеза встречает и другое возражение: развитие язвенного процесса обычно обрывается вместе с элиминацией H. pylori, тогда как однажды индуцированный аутоиммунный процесс должен был бы иметь продолжение. С этой точки зрения значение аутоиммунных реакций выглядит более логичным для атрофического гастрита. В целом H. pylori вызывает широкий спектр реакций иммунитета — от элементов острого (нейтрофилзависимого) воспаления до местных и системных реакций гуморального и клеточного иммунитета. Они недостаточны для элиминации H. pylori, но контролируют его взаимоотношения с хозяином, и, возможно, участвуют в эволюции патологического процесса. Различные факторы не исключают друг друга, но могут действовать вместе как уникальный патогенетический каскад, одним из пусковых механизмов которого служит хеликобактерная инфекция (рис. 2).

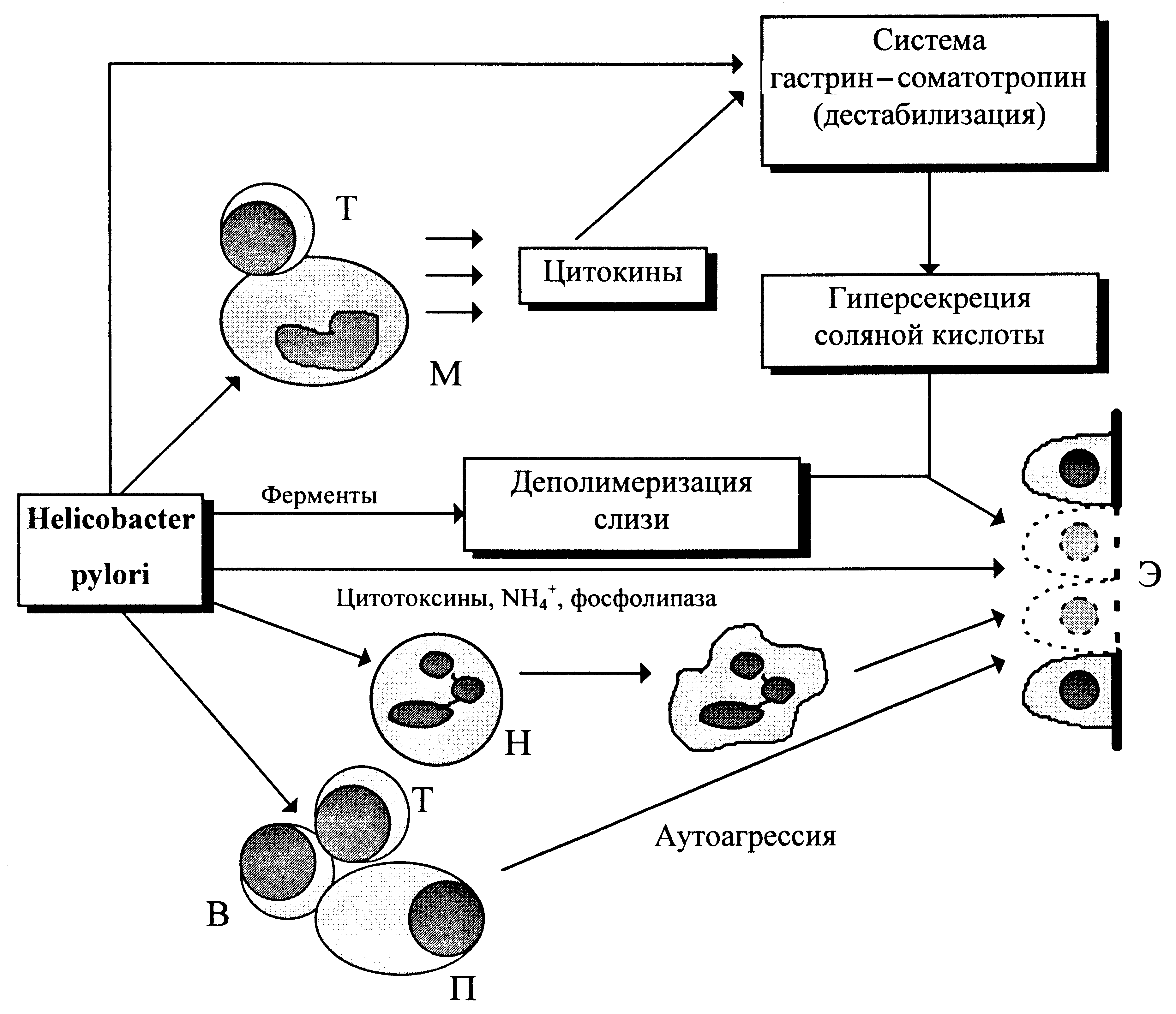
О присутствии H. pylori в желудке (12-перстной кишке) судят по следующим критериям: 1) получение культуры из эндоскопических биоптатов; 2) микроскопия отпечатков биоптатов слизистой оболочки; 3) определение уреазной активности в биопсийном материале или содержимом желудка; 4) выявление антител к хеликобактерным антигенам; 5) тестирование исследуемого материала на присутствие хеликобактерной ДНК.
Один из главных недостатков эндоскопического исследования — инвазивность. Кроме того, при эндоскопии можно не угадать место для взятия биоптата и получить ложноотрицательный результат. Этих недостатков лишен остроумный способ диагностики хеликобактерной инфекции — респираторный тест с мочевиной. Он основан на неинвазивном выявлении уреазной активности в желудке, единственным источником которой является H. pylori. Обследуемый принимает с пищей мочевину, меченную радиоактивным углеродом. Под влиянием уреазы она разлагается на аммиак и углекислый газ, который всасывается в кровь, выделяясь легкими. Его регистрируют в выдыхаемом воздухе, начиная с первых минут после приема мочевины (рис. 3). Метод высоконадежен, так как отражает суммарную метаболическую активность бактерий, независимо от места колонизации в желудке.

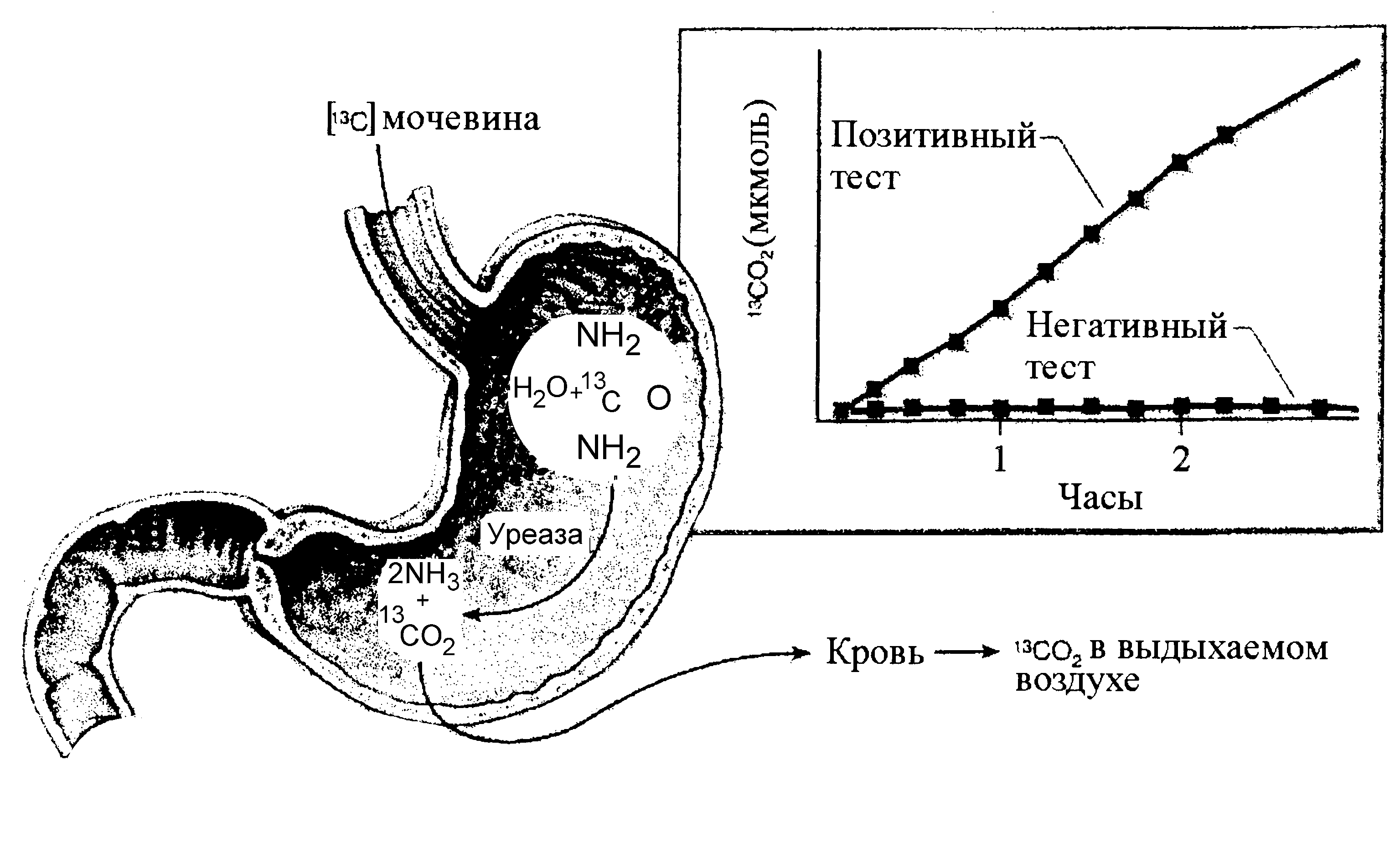
У лиц, инфицированных H. pylori, образуются антитела против различных антигенов H. pylori. Их можно обнаружить в крови и слюне. на этом основана иммунодиагностика хеликобактерной инфекции. Определение сывороточных антител при помощи иммуноферментного анализа по специфичности и чувствительности почти не уступает эндоскопическому обследованию и дыхательной пробе с мочевиной. Такие тесты дешевле и достаточно надежны для выявления инфекции у нелеченых больных. К недостаткам метода относится инерционность: результат остается положительным не менее месяца после ликвидации инфекции. Это затрудняет слежение за эффективностью антибактериальной терапии и полнотой элиминации возбудителя.
Вызывает интерес диагностика хеликобактерной инфекции при помощи полимеразной цепной реакции, которая с предельной чувствительностью позволяет судить о присутствии H. pylori по фрагментам его генома в желудочном соке (в частности генов уреазы — ureA, ureB, ureC).
Открытие хеликобактерной инфекции радикально изменило подходы к лечению больных с язвенной болезнью желудка и 12-перстной кишки: традиционная патогенетическая (антисекреторная) терапия сегодня неизменно сочетается с применением этиотропных (антибактериальных) средств. H. pylori быстро теряет чувствительность к антибиотикам, если они применяются раздельно. В связи с этим предложены разнообразные комбинации и схемы применения антибиотиков и других антибактериальных препаратов, которые во многих случаях позволяют элиминировать H. pylori из организма, создавая условия для выздоровления. Основным требованием является полное искоренение инфекции, о чем в идеале можно судить по негативным результатам микробиологического анализа на протяжении как минимум четырех недель после завершения курса лечения. Впрочем, обычно полагаются на отсутствие рецидивов, и только при их появлении обследование повторяют. Сегодня антибактериальная терапия занимает 1—2 нед, что является большим достижением по сравнению с несколькими месяцами лечения, которое применялось ранее.
Известные надежды связаны с вакцинацией, хотя длительное выживание H. pylori на фоне естественного иммунного ответа заставляет сомневаться в реальности подобной перспективы.
Хеликобактерная инфекция — один из самых популярных объектов современной бактериологии. Интрига сюжета, его актуальность, вызов классическим традициям, противоречия, оставляющие надежды скептикам — все это побуждает к новым изысканиям, которые непрерывно расширяют концептуальные контуры, впервые заложенные чуть более двадцати лет назад. Многое подлежит уточнению, возможны очередные компромиссы. Серьезных последствий, например, следует ожидать от изучения внутривидовой неоднородности H. pylori конституционального и индуцибельного (адаптивного) характера. Это поможет разрешить главное противоречие — бесконфликтность (в том числе иммунологическую) широко распространенного носительства потенциально опасного агента.
Возбудитель сифилиса
Наглому дай волю, он захочет и боле.
Кто знает сифилис, знает медицину.
В. Ослер
- Трепонемы и бледная спирохета.
- Неудачи культивирования.
- Первичная инвазия.
- Клиническая и микробиологическая эволюция процесса.
- Врожденный сифилис.
- Иммунология и иммунопатогенез.
- Парадоксы иммунодиагностики.
- Сальварсан и антибиотики.
Сифилис относится к классическим заболеваниям, передаваемым половым путем. Его изучение занимает заметное место в истории микробиологии, иммунологии и химиотерапии. После недолгого затишья сифилитическая инфекция вновь получила широкое распространение. Ее патогенез остается загадкой. Достоверно известны лишь несколько стадий, которые удивительно многолики и клинически не похожи друг на друга. Две первые из них (первичный и вторичный сифилис) протекают как острое и подострое заболевания, третичный сифилис — хроническая болезнь, которая продолжается много лет и никогда не завершается самоизлечением. Сифилис может заявить о себе в любом возрасте, так как его возбудитель, Treponema pallidum, передается от матери плоду и годами сохраняется в организме, часто в латентном (точнее псевдолатентном) состоянии. Он не культивируется на искусственных питательных средах, и мало известно о факторах, которые определяют его патогенность и способность ускользать от иммунологического надзора. К счастью, T. pallidum сохраняет высокую чувствительность к антибиотикам. Поэтому современный сифилис хорошо поддается лечению, если процесс не достиг необратимых органических поражений.
Название болезни возникло в ХVI столетии, когда в Европе бушевала эпидемия сифилиса. В 1530 г. итальянец Джироламо Фракасторо, много и плодотворно размышлявший о природе заразных болезней, опубликовал поэму, повествующую о судьбе юного пастуха Сифилоса (Syphilos), заболевшего «венерической оспой». Примерно в это же время не менее знаменитый французский врач, математик и астроном Жан Фернель «окрестил» ту же болезнь Lues Venerea, чумой возлюбленных (от лат. lues — чума). С тех пор «сифилис» и «люис» используются как равноправные термины.
Микробиология. В естественных условиях сифилисом болеет только человек. Возбудитель относится к спирохетам — семейству бактерий с уникальной морфологией и структурой. Это спиралевидные, тонкие, гибкие, довольно длинные (до 20 мкм) клетки с активной подвижностью. Спирохеты широко распространены в природе, хотя лишь немногие из них вызывают болезни у человека и животных. Патогенные спирохеты принадлежат к трем родам — Treponema (сифилис, фрамбезия, пинта, беджел), Borrelia (возвратный тиф, болезнь Лайма) и Leptospira (лептоспироз).
Все трепонемы похожи друг на друга, и есть мнение, что они являются вариантами одного вида — T. pallidum, возбудителя сифилиса. Об этом говорит не только их структурное и физиологическое сходство, но и клиническая однотипность трепонемозов. Это хронические инфекции, которые могут продолжаться десятки лет с чередованием обострений и скрытых (латентных) периодов. В отличие от сифилиса, распространенного повсеместно, фрамбезия, пинта и беджел встречаются главным образом в тропических странах и протекают относительно доброкачественно.
Возбудитель сифилиса открыт в 1905 г. Ф. Шаудином и Э. Гоффманом в экссудате тканей, пораженных сифилисом. Он получил название бледной (pallidum) спирохеты, так как, обладая очень тонким телом (ширина клеток 0,1—0,2 мкм), слабо прокрашивается анилиновыми красителями и плохо заметен под обычным микроскопом. Его удается хорошо рассмотреть при помощи темнопольной микроскопии (рис. 1), а также после импрегнации серебром, которое, осаждаясь на клетках, увеличивает их диаметр. В свежеприготовленных темнопольных препаратах трепонемы отличаются характерными движениями: активно перемещаются в среде путем штопорообразного вращения и медленных скачков. Это сопровождается сгибанием клеток под прямым (и более) углом без потери спиралевидной формы. Такого рода сгибательные движения отсутствуют у других спирохет, что является дифференциальным признаком рода Treponema (лат. — сгибающаяся нить).

Строение T. pallidum в принципе не отличается от других спирохет (рис. 2). Цитоплазматический цилиндр окружен плазматической мембраной и плотно прилегающей к нему клеточной стенкой, основу которой составляет пептидогликан — достаточно эластичный, чтобы не мешать сгибательным движениям трепонем. Снаружи есть еще одна оболочка. Она напоминает внешнюю мембрану грамотрицательных бактерий, но в отличие от нее, не содержит липополисахаридного эндотоксина. Между клеточной стенкой и наружной мембраной расположены аксиальные фибриллы, которые плотно обвивают клеточный цилиндр. Они берут начало от базальных дисков, расположенных на обоих концах клетки, и заканчиваются, пройдя середину клеточного цилиндра. По своим свойствам аксиальные фибриллы напоминают жгутики бактерий и поэтому называются эндофлагеллами, т.е. внутренними жгутиками. Полагают, что они обеспечивают подвижность спирохет, хотя это окончательно не доказано. Большинство (50—80%) свежеизолированных штаммов T. pallidum окружены слоем кислых мукополисахаридов, напоминающим капсулу. Однако не все уверены, что это собственный продукт трепонем, а не производное соединительной ткани хозяина. Если это так, то правильнее говорить о псевдокапсуле.

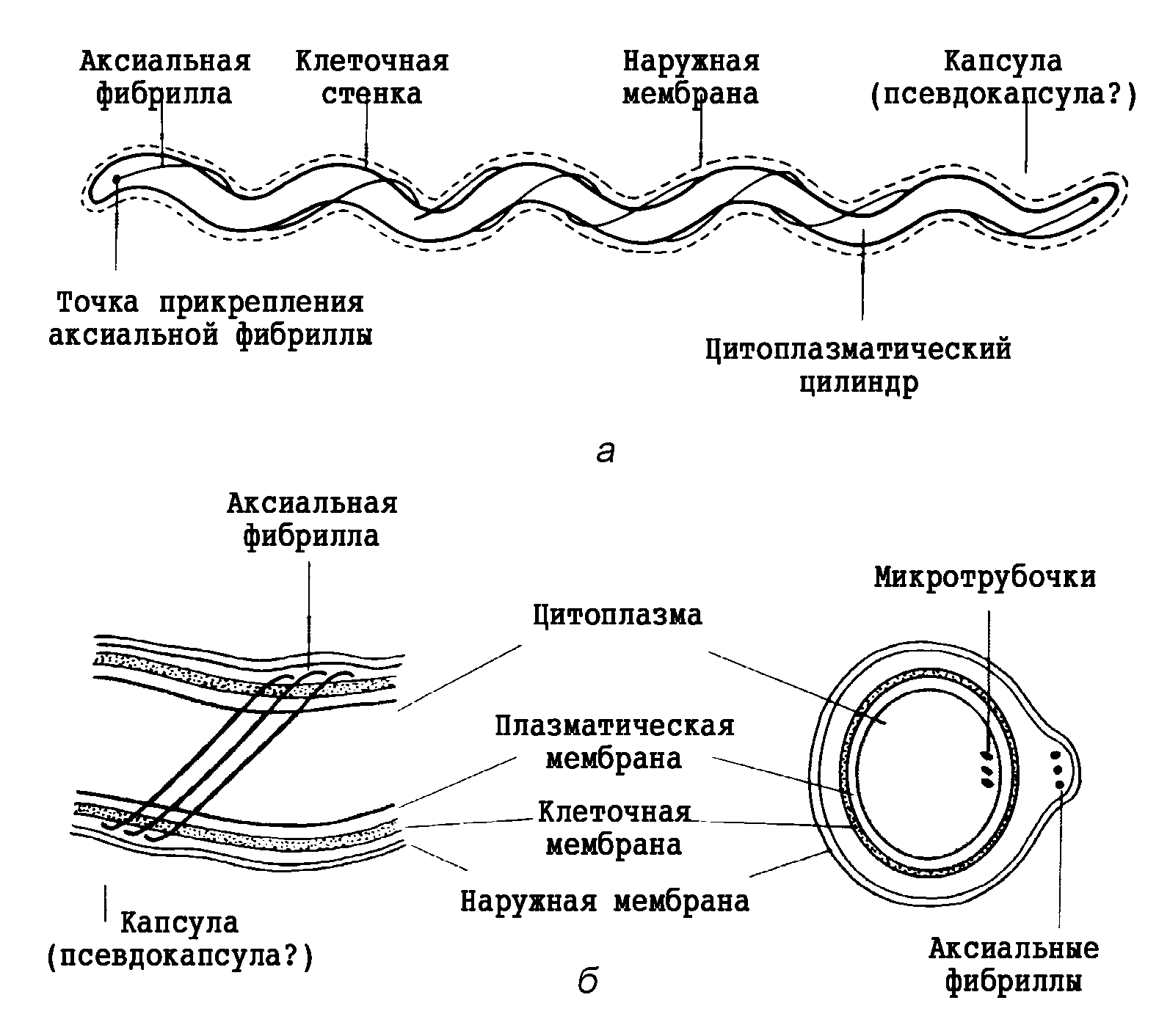
Многочисленные попытки культивировать T. pallidum окончились неудачно. Иногда это как будто удавалось, но при тщательной проверке всегда оказывалось, что вырастал не возбудитель сифилиса, а похожие на него непатогенные трепонемы из нормальной микрофлоры. С появлением клеточных культур возникла надежда, что сифилисная трепонема наконец-то «согласится» реплицироваться in vitro. Однако и в культуре клеток можно поддерживать лишь несколько генераций T. pallidum, получить значительное количество микробной массы не удается. Возбудитель сифилиса отличается необыкновенно медленным для бактерий ростом. Время его генерации составляет около 30 ч, и, возможно, это является барьером для культивирования T. pallidum (для сравнения: обычные бактерии, например кишечная палочка, удваивают число клеток каждые 15 мин). Не следует забывать и о высокой требовательности возбудителя к условиям существования: микроаэрофильность, быстрая гибель при высыхании и повышенной температуре. Было время, когда термолабильность T. pallidum (при 41,5°С они погибают в течение 1 ч) пытались использовать в лечении сифилиса, вызывая у больных приступы лихорадки, например путем заражения малярией.
В связи с невозможностью культивирования решающую роль в изучении T. pallidum сыграли и продолжают играть модели на животных. Первенство принадлежит экспериментальному сифилису кроликов, который был впервые воспроизведен в 1906 г. Кролика удается заразить при помощи любого сифилитического материала, используя различные пути введения: в роговицу, в переднюю камеру глаза, в кожу, в слизистые оболочки и т.д. Наиболее удобным оказалось заражение в паренхиму яичка. Спустя 8—10 дней у животных развивается орхит, а в очаге поражения (первичный сифилис) содержится множество трепонем. Процесс спонтанно разрешается в течение нескольких месяцев, вторичный сифилис развивается редко.
Материал от зараженных кроликов — практически единственный источник трепонем для научных исследований и получения диагностических антигенов. На протяжении многих лет в лабораториях всего мира используется один и тот же штамм T. pallidum — штамм Николса (Nichols), который прошел множество пассажей и отличается высокой вирулентностью. Установлено, что его ИД50 (доза, дающая клинически значимую инфекцию у половины зараженных) составляет всего 57 трепонем для человека и 23 — для кроликов. Кстати, кролики болеют и собственным сифилисом, во многом напоминающим сифилис человека. Его возбудителем является T. paraluiscuniculi, морфологически не отличимая от T. pallidum.
Патология и патогенез. Развитие сифилиса определяется двумя главными механизмами. Один из них (генерализация инфекции) связан с высокой инвазивностью возбудителя. Второй (активация эндогенного инфекта) сопряжен с длительной персистенцией T. pallidum в тканях хозяина, нередко при полном отсутствии клинических симптомов. Склонностью к персистентной инфекции обусловлен тот факт, что микробиологическое выздоровление при сифилисе часто не совпадает с клинической реабилитацией, создавая серьезные диагностические проблемы. На финише (третичный сифилис) в развитие деструктивных реакций включается сам хозяин, который, безуспешно пытаясь освободиться от возбудителя, разряжает эффекторы иммунитета на собственных тканях.
Многолетняя персистенция T. pallidum наводит на мысль о своеобразной коадаптации в системе «хозяин—паразит». Не достигая совершенства, она позволяет микробу периодически выходить из засады, возбуждая клинически многоликий процесс. Элементы коадаптации имеют и популяционные параллели. Первая европейская эпидемия сифилиса, которую связывают с завозом бледной спирохеты из Америки моряками Х. Колумба, имела устрашающий характер, хотя аборигены Нового Света уживались с ней довольно мирно. Адаптивное ослабление вирулентности бледной трепонемы привело к снижению числа катастрофических исходов заболевания, а сам сифилис стал встречаться реже. В 1950—70-х гг. он почти исчез из врачебной практики, создав иллюзию победы, которую поспешили связать с высокой эффективностью антибиотиков. Но к общему разочарованию вскоре возникла новая волна сифилиса, которая вновь вывела его в число лидеров венерической патологии.
Из-за неустойчивости возбудителя во внешней среде заражение сифилисом происходит путем прямого, обычно полового, контакта. Половое общение с инфицированным партнером ведет к заражению примерно в 10% случаев, причем для этого достаточно нескольких десятков трепонем. Внеполовое заражение возможно при тесном контакте с больным (поцелуи) и принадлежащими ему предметами (посуда, бритва, зубная щетка и пр.). Из истории известно немало примеров заражения медицинского персонала при выполнении врачебных манипуляций (например, пальцевый шанкр акушерок). Не менее трагичны и случаи заражения врачами своих пациентов. Сифилис может передаваться при переливании инфицированной крови, хотя с введением специфического контроля за донорским сырьем такая угроза практически отпала, тем более, что опасна лишь свежая кровь; при хранении в холодильнике сифилитическая трепонема погибает через 72—96 ч. Гораздо страшнее и реальнее трансплацентарная передача возбудителя, циркулирующего в крови беременной женщины. В этом случае плод гибнет в утробе матери или родится с врожденным сифилисом, который проявляется в момент рождения или через несколько лет (см. ниже).
Входными воротами для T. pallidum служат слизистые оболочки и кожа. Но возбудитель не задерживается на их поверхности, быстро проникая в ткани через незначительные повреждения. Первые признаки болезни (первичный сифилис) возникают после инкубационного периода длительностью 10—50 дней (в среднем 3—4 нед). В месте внедрения возбудителя (обычно на наружных половых органах) появляется небольшой (1 см в диаметре) плотный безболезненный красноватый узелок, из которого образуется язва — шанкр (от франц. chancre — язва; рис. 3). Она имеет твердое основание и поэтому называется твердым шанкром, в отличие от другой венерической болезни — мягкого шанкра или шанкроида, вызываемого гемофильной палочкой — Haemophilus ducreyi. В отделяемом шанкра обнаруживается множество трепонем, поэтому больной в этом периоде высококонтагиозен.

Одновременно с закреплением и размножением в зоне первичной инвазии трепонемы проникают в лимфатические узлы и вызывают регионарный лимфаденит. Лимфатические узлы увеличиваются до размеров фасоли и становятся плотными, но хорошо подвижными; такие же, но менее выраженные воспалительные изменения развиваются в отдаленных лимфатических узлах (сифилитический лимфаденит). Преодолевая лимфатический барьер, часть трепонем попадает в кровеносное русло, получая возможность для распространения по всему организму. Принципиально, что генерализация инфекции начинается до появления шанкра и лимфаденита, т.е. с самого начала сифилис протекает как системное заболевание, хотя его первой манифестацией может быть единственный очаг поражения кожи или слизистых оболочек.
Первичный сифилис излечивается спонтанно через 2—6 нед, сменяясь временным затишьем, за которым у половины больных спустя 2—24 нед вновь следуют клинические симптомы — вторичный сифилис. Его символом служат поражения кожи и слизистых оболочек (вторичные сифилиды), которые имеют разную локализацию и проявляются в виде полиморфных высыпаний — розеол, папул, пустул и пр. Сифилиды высококонтагиозны, так как содержат множество трепонем. Кроме поверхностных покровов поражаются лимфатические узлы, суставы, кости, внутренние органы, центральная нервная система. В целом клиническая картина может быть столь причудливой, что в прошлом врачи называли вторичный сифилис «великим мистификатором». С микробиологической точки зрения вторичный сифилис следует рассматривать как рецидив эндогенной инфекции, связанный с политропной (полиорганной) репликацией T. pallidum при попустительстве хозяина, который не обеспечивает полноценного иммунитета. Сыпь и другие симптомы бесследно исчезают через несколько недель, но у 25% больных сифилиды рецидивируют на протяжении 2—3 лет.
Примерно треть больных, перенесших вторичный сифилис, спонтанно освобождаются от возбудителя. В остальных случаях формируется бессимптомная инфекция — латентный сифилис, который продолжается от нескольких месяцев до многих лет (иногда пожизненно), в среднем 6—7 лет. У половины носителей латентной инфекции развивается третичный сифилис. Его называют гуммозным, так как для этого периода характерно образование очагов гранулематозного воспаления — гумм (лат. gummi — резина). Их распад приводит к разрушению костных и мягких тканей (например, неба и носа), а также к поражению жизненно важных органов — сердечно-сосудистой системы, печени, спинного и головного мозга. Нарушение кровоснабжения тканей в результате сопутствующего васкулита предрасполагает к инфарктам, способствуя дегенеративным поражениям (в том числе в самих гуммах) с замещением дефектов соединительной тканью. При кардиоваскулярном сифилисе васкулит артерий, питающих торакальную часть аорты, ведет к атрофии ее эластинового слоя и образованию хрупких аневризм; еще одним из типичных осложнений является недостаточность аортального клапана. К тяжелейшим и наиболее поздним осложнениям нейросифилиса относятся спинная сухотка — tabes dorsalis (дегенерация задних столбов, задних корешков и межпозвоночных ганглиев спинного мозга) и прогрессивный паралич (дегенерация нейронов головного мозга), которые развиваются через 5 лет и более с момента заражения (в старых классификациях их даже выделяли в самостоятельный, четвертичный сифилис).
Своевременное применение антибиотиков обрывает естественную эволюцию патологического процесса, и сегодня третичный сифилис — большая редкость. Удивительно, но очаги поражения при третичном сифилисе содержат мало трепонем, а иногда их вообще не удается обнаружить. Поэтому больные в этом периоде неконтагиозны или, как говорят венерологи, третичный сифилис теоретически заразен, а практически — нет.
Врожденный сифилис. Во втором триместре беременности возбудитель, циркулирующий в крови женщины, способен проходить через плаценту и инфицировать плод. Исход заражения определяется числом внедрившихся трепонем, что в свою очередь зависит от стадии сифилитического процесса у матери (следует помнить, что у женщин твердый шанкр часто протекает незаметно). При первичном сифилисе, когда в крови много трепонем, плод может получить большую дозу возбудителя и инфекция завершается внутриутробной смертью (мертворождение) или выкидышем. Если у матери вторичный сифилис или процесс находится в латентной фазе, плод выживает, но развивается врожденный сифилис. Он выявляется при рождении, в течение первых двух лет жизни (ранний врожденный сифилис) или много позже, через 5—20 лет (поздний врожденный сифилис). Считается, что треть беременных женщин, больных сифилисом, имеет здоровое потомство, у другой трети — родятся дети с врожденным сифилисом, в остальных случаях происходят выкидыши или мертворождение.
Симптомы раннего врожденного сифилиса напоминают поражения при вторичном сифилисе взрослых, так как возбудитель сразу попадает в кровь, инициируя системный процесс. Трепонемы в изобилии обнаруживаются в очагах поражения кожи, слизистой оболочки носа, рта, глотки и гортани. Разрушение носовой перегородки вызывает симптом сопения, печень и селезенка увеличены, наблюдаются аномалии длинных костей. Новорожденный похож на старика — сморщенное лицо, дряблая морщинистая кожа, отставание по массе тела, гипотрофия. Поздний врожденный сифилис соответствует третичному периоду. Гуммозные разрушения распространяются на суставы, костную ткань, внутренние органы, половые железы, нервную систему. Характерна триада Гетчинсона: кератит, патология зубов, глухота.
Важно помнить, что врожденный сифилис можно предотвратить. Он возникает только в тех случаях, когда заболевание не диагностируется и не лечится во время беременности. Материнские IgG-антитела проходят через плаценту, но не защищают плод от заражения.
Механизмы и факторы патогенности. Даже краткое знакомство с клиникой сифилиса говорит о своеобразии взаимоотношений возбудителя и хозяина. Инициатива принадлежит трепонемам. Их главным оружием является способность проникать в ткани, сочетая или чередуя локальную и системную инвазию. Это наблюдается с момента инфицирования (лишь часть бактерий остается в зоне первичного инфекта) и продолжается в течение всего процесса. На слизистых оболочках и коже трепонемы не задерживаются, фаза колонизации поверхностных покровов для них не характерна. В тканях T. pallidum размножается вне клеток, возможно, после адгезии на внеклеточном матриксе. Трепонемы располагают рецепторами к фибронектину, ламинину и коллагену, через которые они способны закрепляться в интерстиции. Но такое прикрепление либо обратимо, либо часть трепонем избегает прочной фиксации, активно перемещаясь в межтканевой жидкости и проникая благодаря штопорообразному движению в кровяное русло. Это своеобразный десант, успевающий ускользнуть от воспалительной реакции, которая, хотя и с опозданием (возможно, связанным с медленным размножением трепонем), развивается в виде твердого шанкра. Здесь, как и в других сифилидах, доминируют элементы хронического воспаления, т.е. продуктивно-инфильтративные процессы преобладают над экссудацией. Этим объясняется плотная консистенция сифилитического шанкра.
Трепонемы не продуцируют токсинов, на которые можно было бы возложить ответственность за множественную гибель эпителиальных клеток и изъязвление в очаге первичного сифилиса. Скорее это результат облитерации микрососудов, связанной с васкулитом. Она сопутствует любому периоду сифилиса и играет едва ли не главную роль в сифилитических поражениях.
Первичный сифилис всегда заканчивается самоизлечением: уничтожение местных трепонем ведет к рубцеванию твердого шанкра. Но обычно это временная победа. Укрывшись в тылу хозяина, возбудитель готовит новую, массированную атаку — вторичный сифилис. Парадокс, но механизмы, которые только что успешно справились с первичным процессом и, казалось, должны были бы укрепиться в ходе этой борьбы, не оказывают сопротивления эндогенному рецидиву, протекающему в виде системной инфекции с множеством локальных поражений.
Каким образом возбудителю удается не только выжить, но даже повысить свою агрессивность на фоне нарастающего иммунного ответа? Этот и другие вопросы остаются нерешенными. Высказывалось предположение, что трепонемы выживают, превращаясь в неуязвимые цисты и L-формы. Однако ни то, ни другое не подтвердилось. Цисты в конце концов были низведены до лабораторного феномена, который наблюдается в стареющих культурах непатогенных трепонем, а убедиться в реальности L-форм, якобы обеспечивающих персистенцию возбудителя, оказалось еще сложнее: не умея культивировать бактерии, трудно проследить за их реверсией в типичные (классические) формы. Большинство авторов склоняется к тому, что многолетнее ускользание трепонем от иммунологического надзора связано с маскировкой их антигенов и антигенной изменчивостью. Кроме мукополисахаридов (псевдокапсула?!) T. pallidum покрывается фибронектином и другими белками, оставаясь вне поля зрения эффекторов иммунитета. Не исключено, что, подобно иным возбудителям хронических инфекций (микобактерии, бруцеллы и пр.), трепонемы прячутся внутри клеток хозяина, например в макрофагах. Обсуждается и вопрос о микробзависимой супрессии иммунитета, признаки которой отмечаются у больных вторичным и третичным сифилисом. Это совпадает с присутствием в их крови иммуносупрессивного фактора (возможно, капсульного мукополисахарида T. pallidum), подавляющего ответ Т-лимфоцитов на трепонемные антигены. Возможно и то, что возбудитель сифилиса устойчив к литическому действию комплемента. Это повышает его выживаемость в кровяном русле даже в присутствии антител.
Деструктивные события при третичном сифилисе еще труднее спроецировать на прямую патогенность возбудителя, так как в этом случае очаги поражения содержат мало трепонем. Логичнее поверить в иммунологически опосредованный механизм повреждений, сопряженный с аллергическими реакциями на микробные антигены или с аутоиммунной агрессией против антигенных эпитопов, общих для трепонем и хозяина. Именно к этому моменту формируется гиперчувствительность замедленного типа к трепонемам, которая может быть одной из причин развития гумм — своеобразных очагов хронического воспаления. Впрочем, и такая трактовка разделяется не всеми, хотя развитие аутоиммунных реакций (они проявляются в образовании антител, перекрестно реагирующих с фосфолипидами T. pallidum и тканей хозяина) относится к классике инфекционной иммунологии и служит основой для диагностики сифилиса (см. ниже).
Иммунология. Сифилис принадлежит к инфекциям, хроническое течение которых вступает в несоответствие с выраженным иммунным ответом на антигены возбудителя. Иными словами, понятия «иммунитет» (т.е. резистентность к инфекции) и «иммунные сдвиги» при сифилисе не совпадают, а иногда и противоречат друг другу.
Тот факт, что более чем у половины больных заболевание не выходит за рамки первичной фазы, говорит о том, что иммунитет при сифилисе может быть достаточно эффективен. Символично и то, что у лиц, перенесших вторичный сифилис, повторные заболевания наблюдаются редко. Вместе с тем, во многих случаях иммунологический ответ не предотвращает поэтапного развития инфекции, которая, начавшись с местного процесса, распространяется на весь организм и продолжается много лет.
Иммунологический парадокс сифилиса породил множество экспериментов на животных, главным образом на кроликах. Сложились две точки зрения. Согласно одной из них, устойчивость к реинфекции складывается по типу нестерильного иммунитета, т.е. существует лишь тогда, когда возбудитель сифилиса сохраняется в организме. По другой версии, иммунитет остается и после удаления T. pallidum, т.е. существует независимо от персистентной инфекции. Большинство исследователей склоняется ко второму заключению, но с оговоркой — иммунитет при сифилисе формируется медленно. В этом отношении показательны опыты на кроликах. Раннее (в первые несколько недель) подавление инфекции пенициллином исключает развитие устойчивости к новому заражению. Иммунитет возникает только в том случае, если процесс беспрепятственно развивается 6—8 мес.
Кроме антител при сифилисе формируется клеточный иммунитет, признаки которого выявляются у больных вторичным, латентным и третичным сифилисом в реакциях in vivo (кожные пробы) и in vitro (стимуляция Т-лимфоцитов трепонемными антигенами). Однако роль эффекторных Т-лимфоцитов, как и антител, при сифилисе не ясна. По крайней мере, в опытах на кроликах введение антител или сенсибилизированных лимфоцитов оказывает лишь частичный протективный эффект. Возможно, иммунитет зависит от совместного действия гуморальных и клеточных факторов. Следует помнить и о том, что при третичном сифилисе Т-лимфоциты включаются в развитие гуммозных поражений, т.е. играют патогенетическую, а не защитную роль.
В целом рассуждения об иммунитете при сифилисе затрагивают две проблемы. Содержание одной из них соответствует классической формуле инфекционной иммунологии — приобретению антигениндуцированной устойчивости к экзогенному реинфицированию. Для сифилиса это вероятно, но для реализации такой возможности необходим длительный контакт с возбудителем, чему препятствует раннее прерывание инфекции антибиотиками. Другой вопрос связан с сопротивлением эндогенной инфекции, которая при сифилисе надолго (нередко пожизненно) закрепляется в организме. Почти каноническое мнение о неэффективности иммунитета при сифилисе относится к неспособности хозяина уничтожить персистентную инфекцию, которая определяет эволюцию патологического процесса. Некоторые из причин ускользания трепонем от эффекторов иммунитета прокомментированы выше, но убедительных объяснений длительного сохранения T. pallidum в организме до сих пор не получено.
Лабораторная диагностика. При первичном и вторичном сифилисе T. pallidum можно обнаружить в очагах поражения. Используют микроскопию в темном поле, выявляя нативные (живые) трепонемы, или исследуют фиксированные препараты, применяя иммунофлюоресцентную микроскопию. В обоих случаях необходимо помнить о непатогенных трепонемах, которые входят в состав нормальной микрофлоры и могут присутствовать в исследуемом материале. В темном поле их отличают по морфологии, а при иммунофлюоресценции полагаются на антигенные различия с T.pallidum. При окраске по Граму T. pallidum не выявляются.
Но главным способом лабораторной диагностики сифилиса являются серологические тесты. В латентном периоде это единственная возможность выявления инфекции. Трудно назвать другое заболевание, при котором серологическая диагностика была бы столь разнообразной и имела бы такие же богатые традиции. В 1906 г. Вассерман, Нейссер и Брук предложили первый серологический тест для диагностики сифилиса, ставший классикой прикладной иммунологии. Не умея культивировать T. pallidum, авторы использовали в качестве антигена водный экстракт печени детей, погибших от врожденного сифилиса. Этот материал (инфицированная печень) содержал множество трепонем, и было логичным ожидать, что трепонемные антигены при экстракции перейдут в раствор.
Действительно, в смесях печеночного экстракта и сыворотки больных сифилисом происходило связывание комплемента, что, как известно, отражает образование комплексов «антиген—антитело». Именно такой вывод сделали Вассерман и его коллеги, а изобретенный ими метод (реакция Вассермана) тотчас получил широкое распространение как специфический тест на антитела против возбудителя сифилиса. Но дело оказалось сложнее. Выяснилось, что для получения «сифилисного антигена» вовсе необязательно экстрагировать материал от больных сифилисом: с равным успехом можно использовать печень и другие органы здоровых людей. Пришлось отказаться от ортодоксальной логики, заменив ее неожиданным для современников Вассермана парадоксом: у больных сифилисом появляются антитела против собственных антигенов хозяина! Сифилис — первая патология, при которой были обнаружены аутоантитела, т.е. вскрыт один из механизмов аутоиммунного ответа.
Активный фактор (антиген Вассермана) был идентифицирован в 1941 г. как один из фосфолипидов, входящих в состав митохондриальных мембран. Он известен под названием «кардиолипин», так как после многочисленных испытаний вассермановский реагент стали готовить из бычьего миокарда — ткани, богатой митохондриями. Кардиолипин широко распространен в живой природе, и в конце концов его удалось обнаружить у трепонем. Это побудило к очередному переосмысливанию феномена. Продукция «антител Вассермана» запускается кардиолипином трепонем, который в свободном состоянии лишен иммуногенности, но обретает ее в составе бактерий. Благодаря антигенной общности с тканевыми фосфолипидами такие антитела реагируют с тканевым (митохондриальным) кардиолипином. Этим, в частности, объясняется то, что эпитопный спектр истинных аутоантител против кардиолипина отличается от антикардиолипиновых антител при сифилисе. Такие антитела обнаруживаются при так называемом антифосфолипидном синдроме (например, при системной красной волчанке) и вызывают ложноположительные реакции Вассермана.
Несмотря на эти факты, антикардиолипиновые антитела традиционно называют неспецифическими, нетрепонемными, или (вовсе неудачно) — реагинами (не путать с реагинами немедленной аллергии!), а основанные на них серологические тесты классифицируют как неспецифические, нетрепонемные. Правильнее называть их кардиолипиновыми, помня о том, что они регистрируют антитела против неспецифических (нетрепонемных) фосфолипидов, имеющих общие эпитопы с кардиолипином трепонем. Антикардиолипиновые антитела не связываются с интактными трепонемами, так как фосфолипиды входят в состав плазматической мембраны, экранированной внешними структурами бактериальных клеток.
Вскоре после открытия комплементсвязывающего теста было установлено, что реакцию между вассермановским антигеном и сифилитическими сыворотками можно обнаружить по преципитации (флокуляции). Это послужило основой для разработки множества технических модификаций флокуляционного кардиолипинового теста на сифилис. Благодаря простоте, стандартности, дешевизне и скорости они вытеснили классическую реакцию Вассермана и широко используются для скрининга сифилисной инфекции. В микроварианте (реакция на предметном стекле с оценкой преципитации под микроскопом) результат учитывается через 2—3 мин (рис. 4). Россия — единственная страна, где инерция оказалась сильнее разумного прагматизма: реакция связывания комплемента (между кардиолипином и трепонемными антигенами) остается узаконенным методом лабораторной диагностики сифилиса.


Кардиолипиновые тесты имеют принципиальный недостаток: они не отличаются абсолютной специфичностью для сифилиса. Антитела к кардиолипину выявляются у 0,3—0,9% здоровых людей. Вероятность их обнаружения возрастает при острых инфекционных заболеваниях, но в этом случае они исчезают через несколько недель или месяцев после болезни (острые или транзиторные ложноположительные реакции). Хронические ложноположительные реакции (длительностью более 6 мес) наблюдаются у больных коллагенозами (системная красная волчанка, ревматоидный артрит), проказой, гемолитической анемией, а также у наркоманов и стариков. Полагают, что ложные антикардиолипиновые антитела образуются в ответ на фосфолипиды, высвобождающиеся из поврежденных тканей.
Стремление сделать диагностику более специфичной и учитывать антитела против самого возбудителя, а не только против «чужих» фосфолипидов привело к созданию альтернативного (точнее дополнительного) направления в серодиагностике сифилиса, основанного на выявлении антител против собственных антигенов T. pallidum. Они получили название трепонемных, или специфических тестов. Из-за невозможности культивировать in vitro трепонемы извлекают из тестикулярной ткани кроликов, зараженных штаммом Nichols.
Первой (1948 г.) была разработана реакция иммобилизации трепонем. К живым трепонемам добавляют исследуемую сыворотку и комплемент (сыворотка морской свинки); смесь инкубируют 18 ч при 37оС. В присутствии антител и комплемента трепонемы обездвиживаются и погибают. Метод высокоспецифичен и до сих пор служит эталоном для подтверждения позитивных кардиолипиновых тестов. Его недостаток — техническая сложность и дороговизна, так как живые трепонемы должны поддерживаться путем непрерывных пассажей на кроликах. Этого удается избежать в тестах, основанных на непрямой иммунофлюоресценции. В них используют убитые T. pallidum (штамм Nichols), фиксированные на предметном стекле. Если исследуемая сыворотка содержит специфические антитела, они фиксируются на трепонемах и могут быть обнаружены при помощи меченых антител против иммуноглобулинов человека. Это тоже очень чувствительный и специфичный тест, но и он слишком сложен для повседневной работы. Чаще его используют как референс-контроль для оценки новых методик.
Гораздо доступнее реакции, основанные на антигенах, извлеченных из T. pallidum. Их сорбируют на эритроцитах, выявляя антитела в реакции непрямой гемагглютинации, или фиксируют на полистероловых носителях для иммуноферментного анализа. В настоящее время благодаря генной инженерии появилась возможность клонирования трепонемных генов и получения на их основе рекомбинантных белков. Это открыло новые возможности для изучения иммунных реакций против дискретных антигенов T. pallidum.
Как и при других инфекциях, много внимания уделяется прямому выявлению трепонемных генов при помощи полимеразной цепной реакции. Однако, для того, чтобы использовать этот метод, неоходимо иметь официально зарегистрированные ПЦР-системы.
Динамика антител. В серодиагностике сифилиса пользуются сочетанием как минимум двух тестов, основанных на выявлении антител против кардиолипиновых и трепонемных антигенов. Это дает возможность следить за развитием инфекции, начиная с 7—10-го дня после первых симптомов сифилиса (шанкра), т.е. спустя 3—5 нед после заражения. Целесообразность сочетанного применения кардиолипиновых и трепонемных тестов связана с тем, что динамика их показателей в ходе инфекционного процесса и лечения неодинакова. Так, при первичном сифилисе кардиолипиновые тесты положительны в 60—80% случаев; во вторичном периоде их чувствительность достигает 100%, но затем она постепенно падает, так что около 30% больных третичным сифилисом становятся серонегативными. Антитрепонемные антитела более стабильны, и их обнаружение может быть единственным признаком латентного сифилиса.
Лабильность антикардиолипиновых антител делает их чувствительным индикатором эффективности лечения: если трепонемные тесты долго остаются положительными после успешной терапии, то реакции с кардиолипином обычно угасают через несколько месяцев после элиминации трепонем. Поэтому длительная серопозитивность с высокой вероятностью указывает на сохранение инфекции в организме. Пластичность кардиолипиновых тестов связана с природой антикардиолипиновых антител. В своем большинстве они относятся к IgM, поэтому их синтез не поддерживается клетками иммунологической памяти, прекращаясь после элиминации антигенов. При реинфицировании кардиолипиновые тесты вновь становятся позитивными, отражая реакцию на новое внедрение возбудителя.
В оценке взаимоотношений организма с T. pallidum помогает и дифференцированное выявление антитрепонемных IgM-антител. Обычно они появляются уже через две недели после заражения и быстро (через несколько месяцев) исчезают на фоне адекватной терапии, ведущей к очищению организма от возбудителя. Длительное сохранение IgM-антител позволяет думать, что лечение, несмотря на клиническую ремиссию, микробиологически оказалось неэффективным (возбудитель остался в организме), а положительный результат после серии негативных IgM-тестов говорит о реинфекции. Важное значение имеет определение IgM-антител при врожденной инфекции. Так как IgM не проходит через плаценту, выявление антитрепонемных IgM-антител у новорожденного указывает на врожденный сифилис. Точно также IgM-антитела не проникают через гемато-энцефалический барьер, и их появление в спинномозговой жидкости говорит о нейросифилисе.
Принципы лечения. Сифилис — первая инфекция, в лечении которой был успешно использован синтетический препарат направленного (антиспирохетозного) действия. Это произошло в 1909 г., когда П. Эрлих объявил о сальварсане (лат. спасающий), органическом производном мышьяка, которое обрывало развитие сифилиса у кроликов и излечивало его у человека. В лаборатории Эрлиха сальварсан называли препаратом «606», так как предыдущие синтезированные 605 веществ оказались неэффективными. Сальварсан и его модификации применялись в течение нескольких десятилетий. Однако у них был существенный недостаток — необходимость длительного применения. Это воскрешало афоризм прошлого, когда сифилис пытались лечить ртутными препаратами: «Ночь — с Венерой, оставшуюся жизнь — с Меркурием» (греч. меркурий — ртуть). Кроме того, лечение часто давало осложнения, омрачая мечту П. Эрлиха о «волшебной пуле», которая, убивая паразита, была бы безвредной для хозяина.
Новая эпоха в лечении сифилиса началась с открытия пенициллина. Впервые он был использован с этой целью в 1943 г., и до сих пор его современные варианты остаются препаратами выбора. T. pallidum, по-видимому, не способны приобретать устойчивость к пенициллину и, несмотря на его многолетнее применение, сохраняют высокую чувствительность к антибиотикам этого класса. Эффект пенициллиновой терапии зависит от стадии заболевания и в целом лучше выражен на ранних этапах болезни. При запоздалом лечении дозировки должны быть выше, а курс лечения — длительнее. Известно, что пенициллин действует на растущие бактериальные клетки, блокируя синтез пептидогликана клеточной стенки. Из-за медленного размножения спирохет надежность антитрепонемного эффекта зависит от длительности сохранения адекватной концентрации антибиотика в крови. Этого легче всего добиться при помощи депонированных пенициллинов с пролонгированным действием. В противном случае требуются многократные инъекции препарата. При непереносимости вместо пенициллина используют другие антибиотики, близкие по эффективности (макролиды, тетрациклины, цефалоспорины).
Микоплазмы
Ни рыба, ни мясо, ни кафтан, ни ряса.
- Класс молликут: своеобразие относительно других бактерий.
- Мембранные паразиты.
- Микоплазмы человека: отношение к нормальной микрофлоре и патологии.
- Микоплазменная пневмония и ее возбудитель.
- Патогенность генитальных микоплазм.
Микоплазмы представляют собой мельчайшие бактерии, лишенные ригидной кле точной стенки. Это связано с тем, что у них нет пептидогликана (обязательного атрибута подавляющего большинства бактерий) и его заменителей, стабилизирующих форму клеток. Вместо этого они располагают трехслойной липидной мембраной, которая поддерживает осмотическую прочность микоплазм, но не обеспечивает их постоянной морфологии, особенно во время роста и репликации. Кроме того, многие микоплазмы имеют микрокапсулу, химическая структура которой варьирует у разных видов, но обычно является углеводным полимером.
Микоплазмы размножаются бинарным делением, но образование равновеликих дочерних форм завуалировано из-за десинхронизации роста и деления клеток. Некоторые из них до начала деления успевают многократно реплицировать свой геном (ДНК), образуя удлиненные клетки, которые, вырастая «в сторону», дают ветвящиеся нити (рис. 1). Этим определяется выраженный плеоморфизм микоплазм — от кокковидных до нитевидных, мицелиальных и иных причудливых форм (рис. 2). Самые мелкие из них (элементарные тела), подобно вирусам, проходят сквозь бактериальные фильтры, чему способствует и эластичность клеточной оболочки.

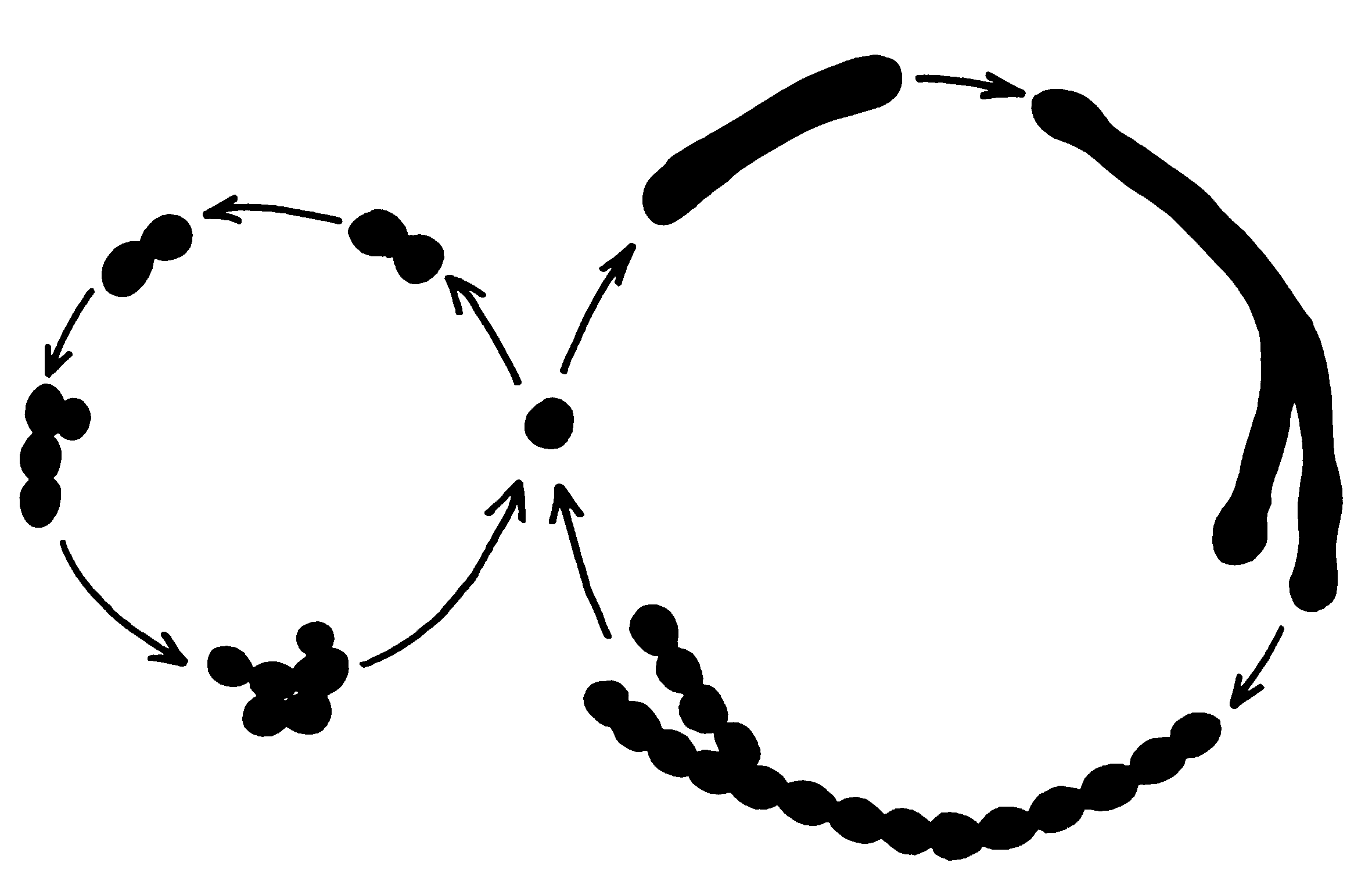

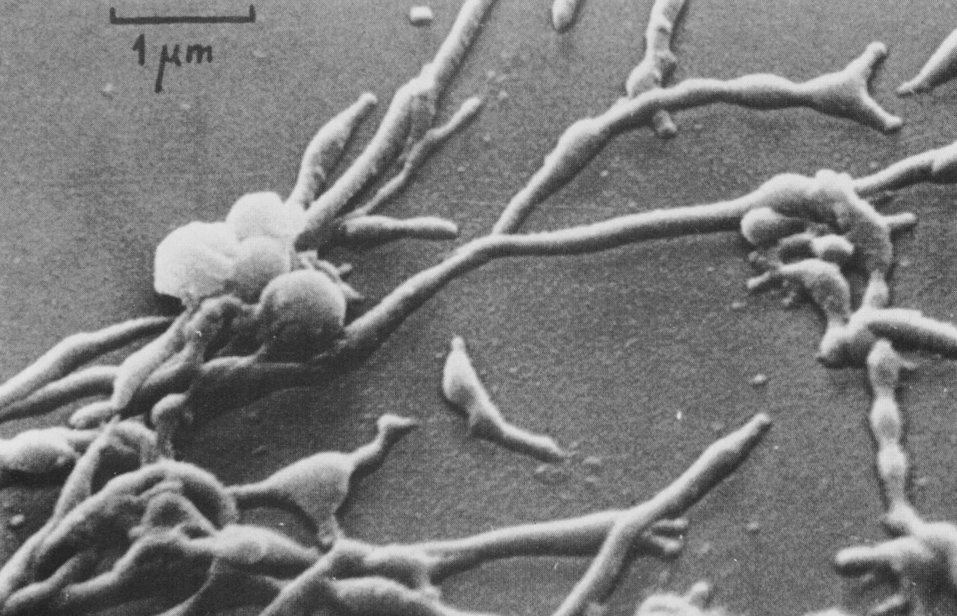
Отсутствие ригидной клеточной стенки закреплено в таксономике микоплазм: они отлучены от остальных прокариот на уровне класса — Mollicutes (лат. мягкокожие). Отношение молликут к бактериям подтверждено наличием общих базисных признаков. Вместе с тем особенности микоплазм столь выразительны, что о них (наряду с хламидиями, риккетсиями, актиномицетами и спирохетами) принято говорить как об «атипичных» бактериях.
Понятие «микоплазмы» сегодня сужено до одного из трех порядков класса Mollicutes — Mycoplаsmatales (единственная группа молликут, представители которой имеют медицинское значение), хотя по инерции ко всем молликутам продолжают обращаться как к микоплазмам. Термин «микоплазмы» формален, отражая способность к образованию грибоподобных мицелиальных форм, типичных для плесневых грибов и актиномицетов (лат. mycos — гриб). Ничего общего с актиномицетами, а тем более с грибами молликуты/микоплазмы не имеют.
Микоплазмы широко распространены в природе. Они паразитируют на человеке, млекопитающих, птицах, насекомых и растениях. Есть виды, которые ведут свободный (сапрофитический) образ жизни в почве и воде. Располагая минимальным для клеток количеством генетического материала (ДНК микоплазм примерно в пять раз меньше, чем у кишечной палочки), микоплазмы обладают ограниченными биосинтетическими возможностями. Поэтому их приходится культивировать на питательных средах, обогащенных предшественниками нуклеиновых кислот, белков и липидов. Но и в оптимальных условиях микоплазмы растут медленно. Колонии на агаровых средах (они имеют плотный, врастающий в среду центр, напоминая яичницу-глазунью — рис. 3) можно обнаружить лишь через 2—3 дня, и даже спустя неделю их размер не превышает 600 мкм, а для некоторых видов (уреаплазмы) — 15—60 мкм в диаметре.

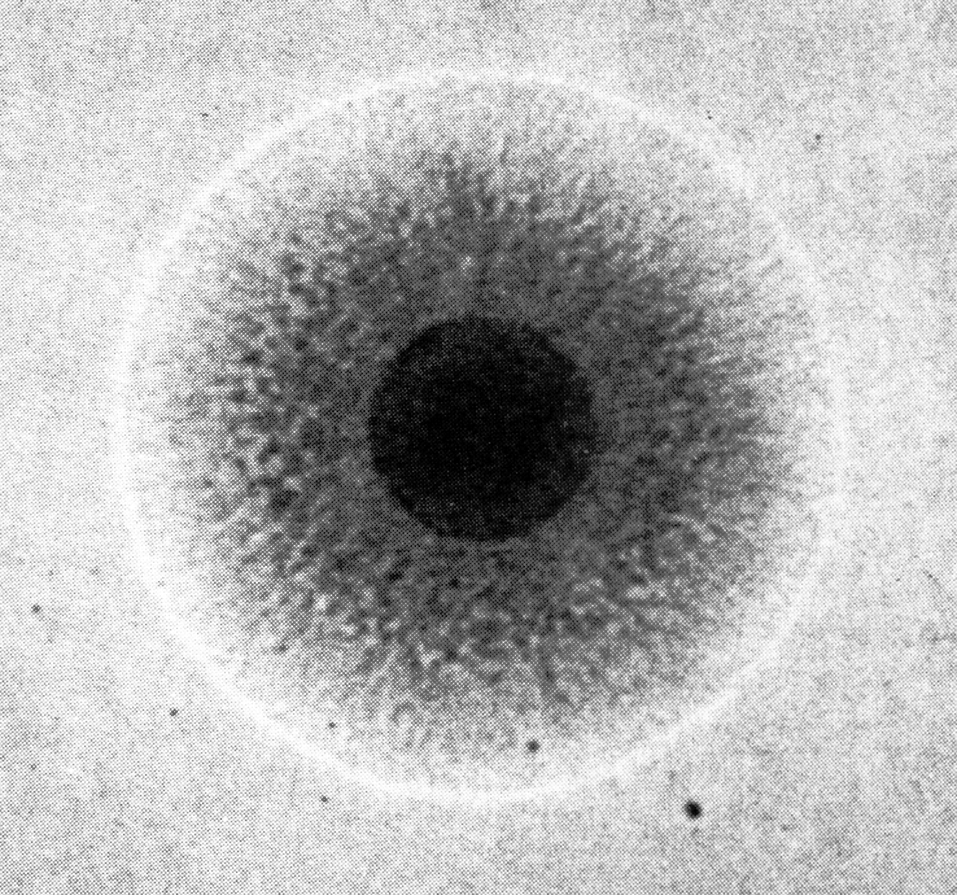
Особенно характерна зависимость микоплазм от стеролов (холестерина и его производных) и жирных кислот. Потребность в холестерине уникальна для бактерий, мембраны которых в отличие от эукариотических клеток обычно лишены стеролов. У микоплазм холестерин доминирует среди липидов, стабилизируя плазматическую и ограничивающую мембраны. Лишь ахолеплазмы (представители одного из порядков молликут) способны обходиться без готового холестерина. В инфицированных тканях микоплазмы добывают стеролы непосредственно из клеточных мембран, внедряясь в их структуру или даже сливаясь с ними. Это оправдывает отношение к микоплазмам как к мембранным паразитам. Что касается энергии, то большинство микоплазм получают ее обычным для факультативных анаэробов способом, ферментируя углеводы или аминокислоты. Облигатно-анаэробные молликуты выделены в самостоятельный порядок (Anaeroplasmatales) и не имеют отношения к медицинской микробиологии.
С человеком связано около десятка видов микоплазм. За двумя исключениями (Ureaplasma urealyticum и Acholeplasma laidlawii) они относятся к роду Mycoplasma. Это неслучайно, так как именно род Mycoplasma представлен наибольшим среди молликут числом видов — около 100; для сравнения уреаплазмы (они названы так из-за уреазной активности, т.е. способности гидролизовать мочевину) включают всего 4 вида. Болезнетворность безоговорочно признается для единственного вида — M. pneumoniae. Еще несколько микоплазм (прежде всего M. hominis и U. urealyticum) претендуют на вклад в патологию человека. Остальные, скорее всего, являются безвредными комменсалами слизистых оболочек (табл. 1).
Таблица 1.
Микоплазмы человека
| Вид | Обычная локализация | Роль в патологии |
| М. pneumoniae | Респираторный тракт | (!) «Атипичная» пневмония |
| М. hominis | Урогенитальный тракт, ротоглотка | (?!) Урогенитальный микоплазмоз |
| М. genitalium | Урогенитальный, респираторный тракты | (?) Урогенитальный микоплазмоз, осложнения микоплазменной пневмонии |
| М. fermentans | Урогенитальный тракт | – |
| М. salivarium | Ротоглотка, гингивальные карманы | (?) Периодонтит |
| М. orale | Верхние дыхательные пути | – |
| М. arthritidis | Синовиальная жидкость (редкий изолят) | (?) Ревматоидный артрит |
| М. incognitus | Различные ткани больных СПИДом (редкий изолят) | (?) Оппортунистические инфекции у больных СПИДом |
| Ureaplasma urealyticum | Урогенитальный тракт | (?!) Урогенитальный микоплазмоз |
| Acholeplasma laidlawii | Ротовая полость, респираторный и урогенитальный тракты |
Микоплазменная пневмония
Респираторный микоплазмоз впервые привлек серьезное внимание в годы второй мировой войны, когда американские врачи отметили вспышки необычной пневмонии среди новобранцев, в казармах (табл. 2).
Таблица 2.
Симптомы микоплазменной пневмонии
| Симптом | Процент больных |
| Кашель | 99 |
| Температура тела >37,8оС | 94 |
| Температура тела >38,9оС | 77 |
| Семья из 4 и более человек | 91 |
| Слабость | 89 |
| Головная боль | 66 |
| Озноб | 58 |
| Фарингит | 54 |
| Мокрота | 45 |
| Хриплость | 37 |
| Ушная боль (субъективно) | 31 |
| Отит (объективно) | 21 |
| Тошнота и/или рвота | 29 |
| Ринит | 29 |
| Лейкоцитоз выше 10 000/мм3 | 27 |
| Лейкоцитоз выше 15 000/мм3 | 5 |
| Рецидивы пневмонии | 22 |
| Сыпь (экзантема) | 17 |
| Диарея | 15 |
| Плеврит | 2 |
| Госпитализация | 2 |
Часто процесс ограничивался острым катаром верхних дыхательных путей (фарингит, ринофарингит, ларингофарингит) и бронхитом, реже поражались глубокие отделы респираторного тракта — первичная атипичная пневмония. Термин означает, что, во-первых, болели здоровые люди («первичная»), а, во-вторых, что их болезнь, в отличие от классической пневмококковой пневмонии, не поддавалась лечению пенициллином и сульфаниламидными препаратами («атипичная»). Интоксикация (температура 38—39°С, слабость, миалгия, артралгии) была умеренной, симптомы нарастали медленно, нередко больные переносили заболевание на ногах (отсюда англ. walking pneumonia — «ходячая пневмония»). Характерным признаком был приступообразный сухой (малопродуктивный) кашель. Рентгенологические симптомы (бронхопневмония) были выражены сильнее, чем этого можно было ожидать по физикальным данным. В крови регистрировали умеренный лейкоцитоз с относительным лимфоцитозом и незначительное повышение СОЭ. Атипичность проявлялась и в том, что у многих (примерно у половины) больных в крови обнаруживались так называемые холодовые гемагглютинины, которые агглютинировали О-эритроциты при температуре 4°С, но не 37°С. Заболевание обычно протекало доброкачественно, выздоровление наступало через 3—4 нед. Фатальные исходы были единичны.
В дальнейшем было сделано множество наблюдений, детализирующих эпидемиологию и клинику инфекции. Оказалось, что респираторный микоплазмоз встречается повсеместно. Чаще поражаются возрастные группы 5—18 лет (около 20% всех случаев пневмонии в этом возрасте); дети до 5 лет и лица старше 20 лет болеют редко. Инфекция распространяется от человека к человеку воздушно-капельным путем, особенно при тесных контактах в закрытых и полузакрытых коллективах (семья, школа, воинский контингент). Накопилось немало наблюдений о внелегочных осложнениях инфекции (табл. 3).
Таблица 3.
Внелегочные проявления инфекции М. pneumoniae
| Зона поражения | Проявления | Число случаев, % |
| Сердечно-сосудистая система | Миокардит, перикардит | <5 |
| Кожа | Мультиформная эритема, макропапулярная сыпь, узелковая эритема, крапивница и др. | Около 5 |
|
Пищеварительный тракт
|
Анорексия, тошнота, рвота, преходящая диарея | 14—44 |
| Гепатит | ? | |
| Панкреатит | ? | |
|
Урогенитальный тракт
|
Тубоовариальный абсцесс | Редко |
| Острый гломерулонефрит | ? | |
|
Система крови
|
Образование холодовых гемагглютининов | Около 50 |
| Гемолитическая анемия | ? | |
| Тромбоцитопения | ? | |
| Внутрисосудистая коагулопатия | ? | |
| Мышечно-скелетная система | Миалгия, артралгия, полиартрит | 14—45 |
| Нервная система | Менингит, менингоэнцефалит, периферическая нейропатия, полимиелитоподобный синдром, мозжечковая атаксия и др. | 6—7 |
Возбудитель был открыт M. Eaton в 1944 г. и долго фигурировал под названием «агента Итона». Считали, что это вирус, так как он проходил через бактериальные фильтры и не культивировался на обычных питательных средах. Лишь в 1960-х гг., когда возникли понятия «микоплазмы» и «микоплазмозы» (заболевания, вызываемые микоплазмами), агент Итона получил статус одного из видов микоплазм — M. pneumoniae.
Мишенью для М. pneumoniae служат клетки реснитчатого эпителия. Это позволяет возбудителю атаковать респираторный тракт на всем протяжении — от носовых ходов до терминальных бронхов (бронхиол). В опытах на добровольцах установлено, что для наиболее вирулентных штаммов достаточно всего 10—100 бактерий (колониеобразующих единиц), чтобы инициировать инфекционный процесс. Возбудитель внедряется между ресничками, прикрепляясь к мембране эпителиоцитов. Вовлечение альвеол опосредовано через интерстициальное воспаление, инициируемое инфекцией мерцательного эпителия бронхов.
М. pneumoniae располагает комплексом адгезинов, главным среди которых является белок Р1. Анти-Р1-антитела блокируют прикрепление микоплазм к клеткам инвитровых эксплантатов бронхов и респираторного тракта искусственно инфицируемых животных. Зацепившись за сиалированные гликопротеины, микоплазмы внедряются в плазматическую мембрану, чему способствуют заостренные концы (терминальные структуры) филаментозных форм, в состав которых входит белок Р1 (рис. 4). Гарантируя устойчивость к вымыванию с обратным током слизи, прочное закрепление микоплазм на мембране обепечивает доступ к клеточным метаболитам (продуктам питания) и способствует повреждению эпителиоцитов. Как и у других микоплазм, инвазия не идет глубже плазматической мембраны (мембранные паразиты!). Проникновение в субэпителиальную ткань, а тем более в кровяное русло, если и происходит, то в виде редкого исключения.




Рис. 4. M. pneumoniae в органной культуре трахеального кольца хомячка через 72 ч после заражения. На рисунке справа терминальные структуры (показаны стрелками) филаментозных форм микоплазм (M) плотно прилегают к основанию ресничек (С) и микроворсинок (m) эпителиального слоя (Е). На рисунке слева видно, что филаментозные формы микоплазм взаимодействуют с клетками, лишенными ресничек. Электронограмма, полоска соответствует 0,2 мкм
Кроме повреждения, связанного с пенетрацией в плазматическую мембрану и заимствованием у нее стеролов, деструктивный эффект может быть обусловлен мембранными белками микоплазм и такими банальными метаболитами, как перекись водорода и супероксидный радикал, которые из-за плотности клеточно-бактериальных контактов не доступны для антиоксидантов. По крайней мере, именно перекись водорода выступает в роли гемолитического фактора M. pneumoniae. Повреждение плазматической мембраны влечет за собой глубокие изменения клеточного метаболизма. Это проявляется, в частности, в нарушении энергозависимых функций реснитчатого аппарата. Обездвиживание ресничек вызывает подавление мукоцилиарного транспорта, т.е. процесса самоочищения слизистиой оболочки респираторного тракта. Нередко восстановление задерживается на несколько недель после клинического выздоровления.
Но патогенез не исчерпывается прямой деструкцией эпителиоцитов. Важную (возможно, решающую) роль играет иммунологически опосредованное повреждение легких. Характер воспалительного инфильтрата при микоплазменной пневмонии отражает преобладание интерстициальных мононуклеарных реакций, напоминающих гиперчувствительность замедленного типа. Поэтому логично, что в опытах на животных иммунодепрессанты блокируют развитие микоплазменной пневмонии, не снижая или даже повышая вероятность генерализации процесса. О значении иммунопатогенетического (инфекционно-аллергического) компонента позволяет думать и возрастной профиль инфекции. Не исключено, например, что низкая чувствительность к М. pneumoniae детей до 5 лет связана с отсутствием гиперчувствительности к микоплазменным антигенам, которая формируется позже под влиянием повторных субклинических инфекций. Следует отметить и длительный инкубационный период заболевания (2—3 нед), в течение которого может происходить сенсибилизация к микоплазменным антигенам и другие иммунологические процессы. Обсуждается вопрос об аутоиммунных реакциях. Они имеют по меньшей мере одно реальное проявление — образование холодовых (низкоаффинных) IgM-гемагглютининов за счет эпитопного родства М. pneumoniae и I-антигена эритроцитов. Но мембранные белки М. pneumoniae перекрестно реагируют и с другими тканями человека — легкого, печени, мозга. К аутоиммунным реакциям следует отнести и образование антииммуноглобулинов (ревматоидного фактора), которое изредка наблюдается у больных микоплазменной пневмонией.
Нельзя забывать и о циркулирующих иммунных комплексах. Они содержат микоплазменные антигены и обнаруживаются у больных с экстрапульмонарными осложнениями. Отлагаясь на базальных мембранах микрососудов, иммунные комплексы способны индуцировать воспалительное поражение внутренних органов, кожи (различные варианты экзантемы), а также коагулопатию.
Но и это не все. У М. pneumoniae обнаружены белки со свойствами суперантигенов, т.е. факторов, неспецифически стимулирующих лимфоциты. Отсюда ряд иммунотропных эффектов микоплазм может быть опосредован через гиперстимуляцию Т- и В-лимфоцитов.
Экстраполируя наблюдения над микоплазмозами животных с достаточно типичной для них генерализацией процесса, можно было бы думать и об инвазивной природе внелегочных осложнений у человека. Однако несмотря на единичные сообщения о выделении М. pneumoniae из спинномозговой жидкости больных с неврологическими осложнениями и наблюдения случаев генерализации респираторного микоплазмоза у экспериментальных животных, большинство авторов убеждено в противном, полагая, что возбудитель лишен инвазивности и не выходит за пределы респираторного тракта. Не продуцирует он и токсинов дистантного действия. Это подчеркивает важность реактивных поражений, связанных с иммунотропностью M. pneumoniae. Несомненно и то, что эволюция микоплазменного процесса во многом зависит от иммунологической индивидуальности хозяина, которая препятствует либо способствует реализации болезнетворности микоплазм. Не случайно клинические проявления инфекции наблюдаются лишь у 3—10% инфицированных, а такой символичный иммунологический симптом, как холодовые гемагглютинины, обнаруживается лишь у 30—50% больных. В целом респираторный микоплазмоз отражает патогенетическую атипичность микоплазм, которая базируется на необычности их взаимоотношений с хозяином.
Микоплазменная пневмония сопровождается накоплением сывороточных антител, выявляемых в различных тестах (реакция связывания комплемента, преципитация, подавление роста микоплазм, иммуноферментный анализ и др.). Но с учетом низкой инвазивности микоплазм протективная функция циркулирующих антител сомнительна. Скорее всего это антитела-свидетели, которые имеют диагностическое значение, но не участвуют в защите организма от инфекции. Более реально значение IgA-антител респираторных секретов: блокируя микоплазменные адгезины, они обрывают инфекцию в самом начале. Не исключена и роль клеточного иммунитета. На первый взгляд, это идет вразрез с представлениями об иммунологически зависимой болезнетворности микоплазм. Но это кажущееся противоречие. Если иммунопатогенез предполагает массированную стимуляцию Т-лимфоцитов утвердившимся инфектом, то протективное действие основано на опережающем (профилактическом) эффекте сенсибилизированных Т-лимфоцитов, которые уничтожают возбудитель до его накопления в патогенетически значимой дозе.
Учитывая данные о быстром снижении титров антител после респираторного микоплазмоза, можно утверждать, что стабилизация иммунитета к M. pneumoniaе, по-видимому, связана с его подстегиванием при повторных встречах с инфекцией. Этому способствует антигенная однородность M. pneumoniae, которая обеспечивает перекрестный иммунитет едва ли не ко всем штаммам. Но возможны и рецидивы. Кроме того, возбудитель обычно не сразу изгоняется из организма после клинического выздоровления, формируя прослойку бактерионосителей.
Для лабораторного подтверждения респираторного микоплазмоза на практике пользуются определением титра холодовых гемагглютининов, но лучше анализировать антимикоплазменные антитела, например, в реакции связывания комплемента. Таким путем удается повысить диагностическую значимость серологических тестов до 80% (четырехкратное и более повышение титра антител в динамике процесса, или титр 1:64 и более при однократном исследовании). Выделение M. pneumoniae возможно, но требует значительного времени и поэтому проводится редко. Определение микоплазменных антигенов (иммунофлюоресценция, иммуноферментный метод и др.) в мокроте больных ограничено из-за слабой чувствительности и перекрестных реакций с другими микоплазмами респираторного тракта (например, M. genitalium). Большим прогрессом явилось внедрение быстрого и чувствительного определения генов M. pneumoniae в полимеразной цепной реакции (ПЦР). Для получения культуры рекомендуют продолжить изоляцию микоплазм из ПЦР-положительных образцов.
Урогенитальные микоплазмы
Некоторые из микоплазм отдают предпочтение урогенитальному тракту человека, передаваясь половым путем. Их собирательно называют урогенитальными, или просто генитальными микоплазмами. Главными представителями являются два вида — Mycoplasma hominis и Ureaplasma urealyticum. Есть основания, чтобы считать их болезнетворными, хотя до сих пор это делается с оговорками. Сомнения связаны с широким распространением здорового носительства генитальных микоплазм, которое по некоторым данным достигает 50% и более. Вместе с тем анализ иммуносерологических сдвигов (титры и динамика антимикоплазменных антител при патологических процессах), опыты на животных, тестирование в клеточных культурах позволяют думать о патогенетическом потенциале M. hominis и U. urealyticum.
Круг заболеваний, которые с большей или меньшей вероятностью ассоциированы с ними, выглядит устрашающим, особенно для женщин, плодов и новорожденных. M. hominis и U. urealyticum способны инфицировать половой тракт на всем его протяжении — от влагалища до органов малого таза. Их (чаще M. hominis) выделяют при вагините и вагинозе, остром и хроническом сальпингите, сальпингоофорите, тубоовариальных абсцессах, параметрите, эндометрите, аднексите, воспалении тазовой брюшины и клетчатки. Это может вести к бесплодию, осложнять течение беременности (внематочная беременность, преждевременные роды и мертворождение из-за поражения плаценты и околоплодных оболочек), вызывать внутриутробное инфицирование. В последнем случае вероятен генерализованный процесс с поражением органов дыхания, зрения, печени, почек, центральной нервной системы, кожных покровов плода и новорожденных. Это наиболее типично для недоношенных, имевших длительный и обильный контакт с материнскими микоплазмами из-за повреждения околоплодных мембран. Одним из возможных проявлений микоплазменной инфекции у женщин является послеродовая или послеабортная лихорадка: примерно у 10% женщин с септическими послеродовыми симптомами из крови выделяются M. hominis и/или U. urealyticum. Что касается поражений мочевыводящей системы, то для женщин характерно бессимптомное инфицирование уретры и мочевого пузыря, которое чаще наблюдается при беременности и загадочным образом повышает вероятность преждевременных родов и преэклампсии. Чаще выделяется U. urealyticum, что может быть связано с уреазной активностью уреаплазм, а потому с их большей приспособленностью к данному биотопу. Уреаплазмы обладают еще одной особенностью, которая может иметь патогенетическое значение: они продуцируют фосфолипазы А и С, запуская высвобождение арахидоновой кислоты из мембранных фосфолипидов. Не исключено, что, инфицируя амнион, они используют в качестве субстрата его мембраны, способствуя гиперпродукции простагландинов, провоцирующих спонтанные аборты и преждевременные роды. Это может иметь значение и при уреаплазменных поражениях респираторного тракта плода. Заметим, однако, что другие микоплазмы, в частности M. hominis и M. pneumoniae, лишены фосфолипаз, и это не отрицает их патогенности. То же самое следует сказать о протеазе, разрушающей IgA (т.е. антитела слизистых оболочек): она уникальна для U. urealyticum.
У мужчин большинство проблем (часто гипотетичных!), связано с уреаплазменной инфекцией мочевыводящих путей, главным образом уретритом. Это сравнительно немногочисленная группа больных, у которых заболевание не ассоциировано с гонококком и хламидиями. Еще менее очевидна роль микоплазм при восходящей инфекции мочевыводящего тракта и половой сферы (простатит, эпидидимит). Впрочем, подобная вероятность не исключается, как и то, что уреаплазмы инфицируют сперматозоиды, косвенно влияя на процесс репродукции (снижение подвижности и пенетрации сперматозоидов в яйцеклетку).
Нельзя игнорировать и сообщения о выделении M. hominis (включая бактериемию) от больных с хирургическими операциями, из гематом, очагов септического артрита, аспирационной эмпиемы, септического тромбофлебита, перитонита, абсцессов мозга. Эти наблюдения дополняют представления об экстрагенитальных поражениях при врожденных микоплазмозах.
В целом, соблазн возложить на генитальные микоплазмы ответственность за многие «беды человечества» вполне логичен. Надо лишь искать новые принципы, чтобы преодолеть традиционные противоречия, которые со времен Р. Коха тревожили микробиологов и патологов. Главной зацепкой для скептиков служит широкое распространение здорового носительства микоплазм среди взрослых и детей, заразившихся при прохождении инфицированного родового канала. Одним из объяснений может быть внутривидовая неоднородность M. hominis и U. urealyticum, которая отражается на их болезнетворности. По антигенным свойствам уреаплазмы человека делятся на 16 серотипов. Их патогенетические особенности четко не установлены, но есть данные, позволяющие думать об этом. Например, серотип U. urealyticum, выделяемый при патологии новорожденных (младенцы с низкой массой тела), не совпадает с серотипом штаммов, персистирующих в родовом канале их матерей. Различные изоляты M. hominis гетерогенны по рибосомальным РНК, разбросаны по гомологии ДНК, антигенным и культуральным свойствам.
Компромиссом является признание за M. hominis и U. urealyticum статуса оппортунистических патогенов, опасных для лиц с ослаблением местного и общего иммунитета. Это подтверждают наблюдения о микоплазменной бактериемии у больных с агаммаглобулинемией, лимфогранулематозом, почечными трансплантатами, заболеваниями сердца, лейкемией, лимфомами, ревматоидным артритом, тяжелой травмой. В таких случаях микоплазмы (прежде всего M. hominis) способны, по-видимому, диссеминировать с кровью по организму, вызывая экстрагенитальную патологию, например септические артриты, поражения митральных клапанов, аорты и др. В этой связи напомним об ажиотаже (1989 г.) после выделения из саркомы Капоши у больных СПИДом нового вида микоплазм — M. incognitus (лат. таинственный, неизвестный), окрещенного СПИД-ассоциированной микоплазмой. В отличие от других микоплазм человека M. incognitus размножается внутриклеточно (в цитоплазме и ядре инфицированных клеток) и, возможно, действует как дополнительный иммуносупрессивный агент. Но скорее всего это один из спутников ВИЧ-инфекции, ближе всего стоящих к ранее описанному виду микоплазм — M. fermentans.
Чуть раньше был открыт еще один вид, M. genitalium, названный так потому, что он был впервые выделен из генитального тракта гомосексуалистов с негонококковым уретритом. Позже оказалось, что M. genitalium охотно колонизирует и респираторный тракт. Не исключено, что он действует как оппортунист, утяжеляя микоплазменную (M. pneumoniae) пневмонию и способствуя ее экстрапульмонарным осложнениям. Это один из самых труднокультивируемых видов микоплазм, генетически близкий M. pneumoniae. Его привередливость наводит на мысль о существовании других, пока неизвестных, микоплазм, которые могут выступать в роли первичных, но скорее оппортунистических патогенов.
Уместно сказать еще об одной проблеме, препятствующей безоговорочному признанию болезнетворности генитальных микоплазм: в очагах поражения они нередко сосуществуют с микроорганизмами, за которыми утвердилось лидерство в урогенитальной патологии. Их слабая конкурентоспособность с хламидиями, герпесвирусами, а тем более с гонококком очевидна. Но и в таких случаях немало оснований думать об оппортунистической микоплазменной инфекции, которая осложняет основное заболевание, способствуя хронизации воспалительного процесса. Прояснить ситуацию могли бы опыты на добровольцах. Нам известны попытки воспроизвести уретрит при введении в уретру чистой культуры микоплазм: в двух случаях из трех для U. urealiticum получен положительный результат; заражение M. hominis было неудачным.
Практическим выходом при лечении служит то, что, нацеливаясь на уничтожение одного возбудителя, обычно действуют и на другие. Надо лишь правильно выбрать лекарство. Микоплазмы (это столь же справедливо и для хламидий) резистентны к антибиотикам, подавляющим синтез клеточной стенки (ее у них нет), но чувствительны к ингибиторам белкового синтеза. Именно из их числа выбирают препараты для лечения урогенитальных инфекций. Следует только помнить, что по сравнению с M. pneumoniae генитальные микоплазмы более вариабельны по чувствительности к антибиотикам, а устойчивость к тетрациклину и эритромицину распространена достаточно широко.
Что касается специфической диагностики урогенитального микоплазмоза/уреаплазмоза, то, кроме выделения чистой культуры, имеются неплохие возможности для выявления микробных антигенов и нуклеиновых кислот (ПЦР) в очагах поражения. Выявление антител в сыворотке не надежно, так как они часто обнаруживаются у здоровых людей, а острые инфекции (особенно U. urealyticum) не всегда сопровождаются повышением их титра. Впрочем, это справедливо и для определения микробных молекул в исследуемом материале.
Наиболее важный контингент для анализа — беременные женщины, поскольку именно в этом случае возможны тяжелые (но окончательно не доказанные) последствия урогенитального микоплазмоза. Микробиологические признаки микоплазменной инфекции должны быть подкреплены данными об активности патологического (воспалительного) процесса. Только в этом случае стоит задуматься над целесообразностью активной антибактериальной терапии.
Интерес к микоплазмам не ослабевает. Не случайно микоплазмология выросла в самостоятельный раздел бактериологии со своеобразной идеологией и методами исследования. Мы затронули лишь те позиции, которые наиболее убедительно ассоциированы с клинической патологией. Но претензии микоплазмологов гораздо шире. Микоплазмы изучают в связи с проблемами иммунопатологии, онкогенеза, нарушений гемопоэза, системных коллагенозов. Немало внимания уделяется смешанным инфекциям микоплазм с бактериями и вирусами. Будущее покажет, что реальнее — увлечения энтузиастов или сомнения скептиков. Опыт с урогенитальными микоплазмозами показывает возможность компромисса между ними.
Хламидии
У всякой пташки свои замашки.
- Принципы современной классификации.
- Природа внутриклеточного паразитизма.
- Репликативный цикл: взаимопревращения элементарных и ретикулярных тел.
- Механизм цитопатического эффекта.
- Чувствительность к антибиотикам.
- Таксономика: фенотипические и генотипические параллели.
- Ch. trachomatis: патогенетическая многоликость.
- Ch. pneumoniae: реальность и спекуляции.
- Ch. psittaci: пситтакоз-орнитоз.
Хламидии были открыты в 1907 г., когда Л. Гальбштедтер и С. Провацек (эти исследования проводились на острове Ява) сначала экспериментально доказали контагиозность трахомы, заразив орангутангов материалом из соскоба конъюнктивы больных трахомой, а затем в окрашенных клетках пораженной конъюнктивы обнаружили цитоплазматические включения с множеством мельчайших частиц (рис. 1). Они сделали справедливый вывод о том, что это и есть возбудитель трахомы. Ими же было предложено название Chlamydozoa (от греч. chlamys — мантия), которое отражает наличие оболочки вокруг микробных частиц. В измененном виде это название закрепилось в микробиологии, хотя и является по сути ошибочным: оболочка представляет собой продукт клетки-хозяина, а не хламидий.

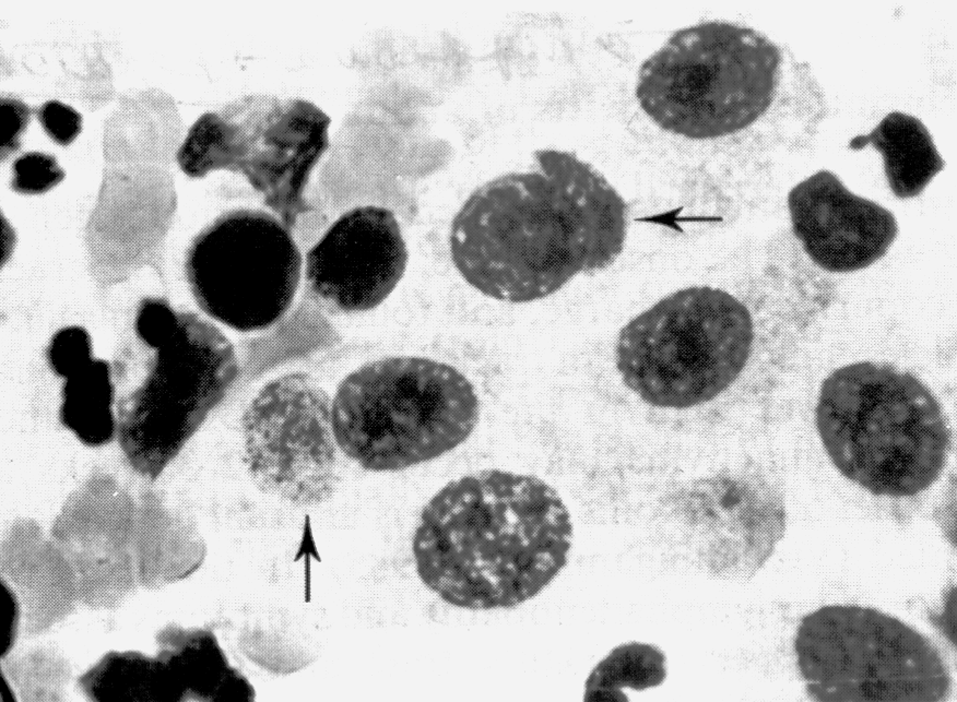
Вскоре такие же включения (тельца Гальбштедтера—Провацека) были обнаружены в конъюнктивальных клетках новорожденных с негонококковой бленореей, в цервикальном эпителии их матерей и в эпителии уретры мужчин с негонорейным уретритом. Стало очевидным, что трахома, конъюнктивит новорожденных с включениями и поражение генитального тракта взрослых вызываются сходными инфекционными агентами. Благодаря способности проходить через фильтры, задерживающие бактерии, а также невозможности культивировать их на искусственных питательных средах, долго поддерживалось ошибочное мнение о вирусной природе хламидий.
В 1930 г. С. Бедсоном был открыт возбудитель пситтакоза и описан цикл его развития. Он высказался о бактериальной природе возбудителя, отметив облигатность его внутриклеточного паразитизма. Немногим позже было обращено внимание на сходство репликативного механизма и морфологии включений возбудителей пситтакоза и группы трахомы. Открытие общего комплементсвязывающего антигена укрепило эту позицию, и они (вместе с возбудителями венерической лимфогранулемы и мышиной пневмонии) были объединены в родовой таксон, который с 1945 г. стали называть Chlamydia.
Современная эра в изучении хламидий началась в 1960-х гг. после разработки методов их выделения сначала в желточном мешке куриного эмбриона, а затем в культуре клеток. К этому периоду относится окончательное утверждение этиологической значимости хламидий в патологии человека (заражение чистыми культурами добровольцев) и установление базисных закономерностей их физиологии. В 1980-х гг. был обнаружен новый вид человеческих хламидий — Сh. pneumoniae, и сегодня ни у кого не вызывает сомнений, что хламидиозы принадлежат к числу наиболее распространенных инфекционных заболеваний человека.
Хламидии представляют собой мелкие сферические или овоидные грамотрицательные бактерии, которые размножаются в цитоплазме живых клеток, проходя через уникальный репликативный цикл. Они не культивируются на искусственных питательных средах, т.е., подобно вирусам, являются облигатными внутриклеточными паразитами. Но это принципиально иной тип паразитизма. Зависимость хламидий от клеток объясняется их неспособностью аккумулировать и использовать энергию, так как они не умеют (или плохо умеют) синтезировать аденозинтрифосфат и грамотно распоряжаться его макроэргическими связями. Это делает за них клетка-хозяин, в связи с чем хламидии называют энергетическими паразитами (о вирусах говорят как о генетических паразитах).
Для хламидий характерен плеоморфизм. Внутри инфицированных клеток они представлены набором кокковидных форм, которые различаются по размеру, тинкториальным свойствам, проницаемости для электронов, плотности клеточной оболочки, соотношению РНК/ДНК и др. Эта неоднородность возникает в ходе репликативного цикла, отражая взаимопревращения двух основных морфофункциональных элементов хламидий — элементарных и ретикулярных тел (рис. 2). Элементарные тела (ЭТ) имеют 0,2—0,3 мкм в диаметре (отсюда вирусоподобная фильтруемость хламидий), а на электронограммах выглядят темными из-за спрессованности их материала в минимальном объеме. ЭТ обеспечивают выживание хламидий во внеклеточной среде и заражение новых клеток, т.е. служат инфекционным началом хламидий. Ретикулярные тела (РТ) — крупнее (0,8—1,5 мкм) и хорошо проницаемы для электронов (отсюда название — ретикулярные). Они вырастают из ЭТ внутри клеток, лишены инфекциозности и, подвергаясь делению, обеспечивают репродукцию хламидий (см. ниже). Отсюда другое, исторически первое название РТ — инициальное тело. Кроме классических ЭТ и РТ внутриклеточные включения содержат промежуточные формы, возникающие при взаимопревращениях ЭТ и РТ.

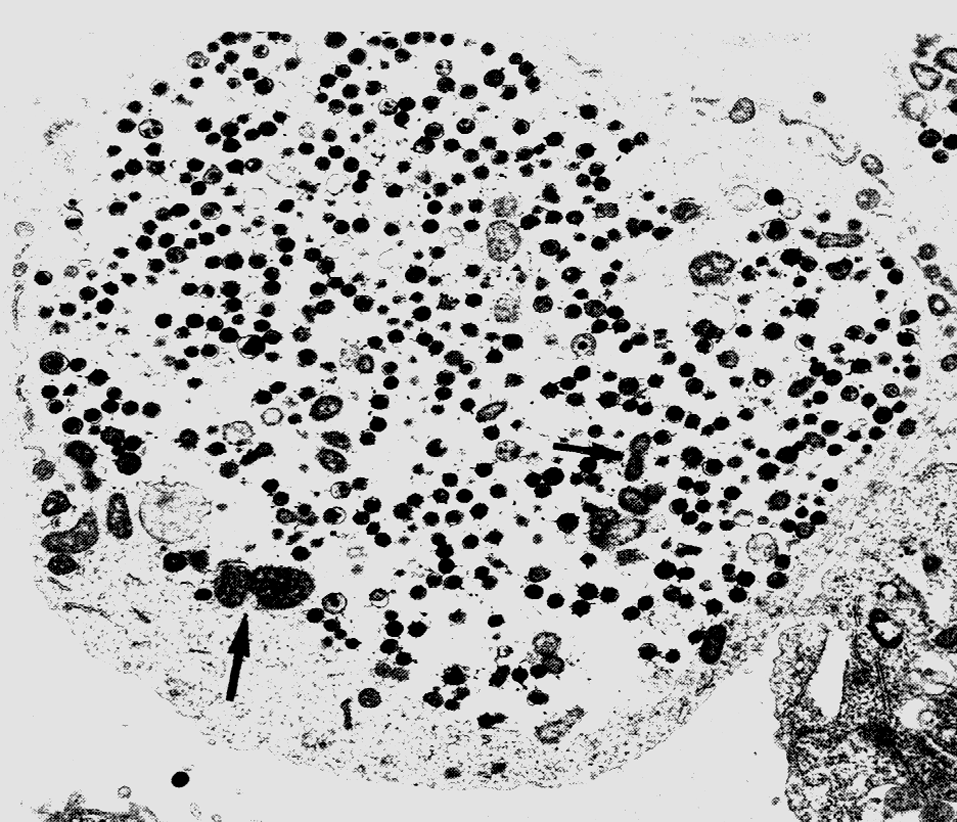
Своеобразие хламидий проявляется и в строении их клеточной стенки. Она лишена пептидогликана и представляет собой двухслойную мембрану, ригидность которой определяют пептиды, перекрестно сшитые дисульфидными мостиками. Умеренная чувствительность хламидий к пенициллину и другим беталактамным антибиотикам объясняется тем, что они препятствуют полимеризации мембранных пептидов, обратимо блокируя образование ЭТ. В остальном хламидии напоминают грамотрицательные бактерии, прежде всего тем, что содержат гликолипиды, сходные с липополисахаридами (ЛПС-эндотоксинами) внешней мембраны грамотрицательных бактерий. Кроме того, хламидии располагают комплексом мембранных белков/пептидов, среди которых доминирует так называемый главный белок наружной мембраны, составляющий около 60% всего белка клеточной стенки. Липополисахариды (гликолипиды) несут эпитопы, общие для всех хламидий (родоспецифический антиген). С главным белком наружной мембраны связано большинство видоспецифических эпитопов (по крайней мере для Ch. trachomatis). Кроме того, Сh. trachomatis и Сh. рsittaci располагают типоспецифическими антигенами, которые определяют внутривидовые особенности хламидий. Скорее всего они относятся к семейству мембранных пептидов, хотя эпитопами могут быть гаптеновые группировки небелковой природы.
Элементарные тела лишены метаболической активности, а плотная наружная мембрана делает их относительно устойчивыми, позволяя переносить невзгоды внеклеточной среды. Сталкиваясь с чувствительными клетками, ЭТ фиксируются на плазматической мембране и индуцируют фагоцитоз. Принципиально, что к этому побуждаются клетки, не способные к активному фагоцитозу, в частности эпителиоциты — главная мишень для хламидий в организме. Об инициативе ЭТ на этом этапе говорит то, что убитые хламидии, а также хламидии, покрытые антителами, не поглощаются эпителиоцитами. Природа хламидийных факторов и клеточных рецепторов, задействованных в этом процессе, остается неясной. Показано лишь, что они отличаются у разных хламидий и клеток и конечный результат зависит от ряда обстоятельств, например от заряда клеточной поверхности. На роль адгезинов претендуют несколько пептидов. Они имеются только на ЭТ, и некоторые из них комплементарны нейраминовой кислоте. Но эта связка не универсальна: она вероятна лишь для «глазной» биогруппы Ch. trachomatis (см. ниже). Есть данные об участии в поглотительных реакциях аппарата секреции III типа.
После поглощения ЭТ оказываются внутри фагосомы. Они вновь ведут себя нестандартно, блокируя слияние фагосомы с лизосомами, по крайней мере до окончания репликативного цикла. Благодаря этому хламидии избегают контакта с антимикробными факторами, получая возможность для беспрепятственного внутриклеточного развития. Оказавшись внутри клеток, ЭТ трансформируются в РТ. Это сопровождается структурной перестройкой мембран, которая повышает проницаемость клеток для АТФ и других питательных веществ. Говоря об уникальности жизненного цикла хламидий, следует помнить и о его известной аналогии со спорообразующими бактериями. С функциональной и экологической точек зрения ЭТ напоминают бактериальные эндоспоры. И те и другие предназначены для выживания во внешней среде, заражения новых хозяев и прорастания в вегетативные (т.е. способные к размножению) клетки. Для хламидий внешняя среда начинается тотчас после высвобождения из клеток. С этого момента стартует «звездный час» ЭТ с их относительной устойчивостью и способностью активно проникать в новые клетки.
Ретикулярные тела являются вегетативной формой хламидий. Отсутствие плотной оболочки делает их более хрупкими, чем ЭТ, но это и пробуждает в них жизнь, обеспечивая метаболические контакты с клеткой-хозяином. РТ многократно делятся, а затем (через промежуточные формы) превращаются в ЭТ. Это совершается не одновременно, а потому на протяжении всего цикла внутри клеток можно видеть РТ, ЭТ и промежуточные формы (см. рис. 2). При множественном заражении образуется несколько фагосом — вакуолей, содержащих ЭТ. В дальнейшем они сливаются вместе (Сh. trachomatis; см. рис. 1) либо дают начало дискретным внутриклеточным включениям (Ch. psittaci; рис. 3).

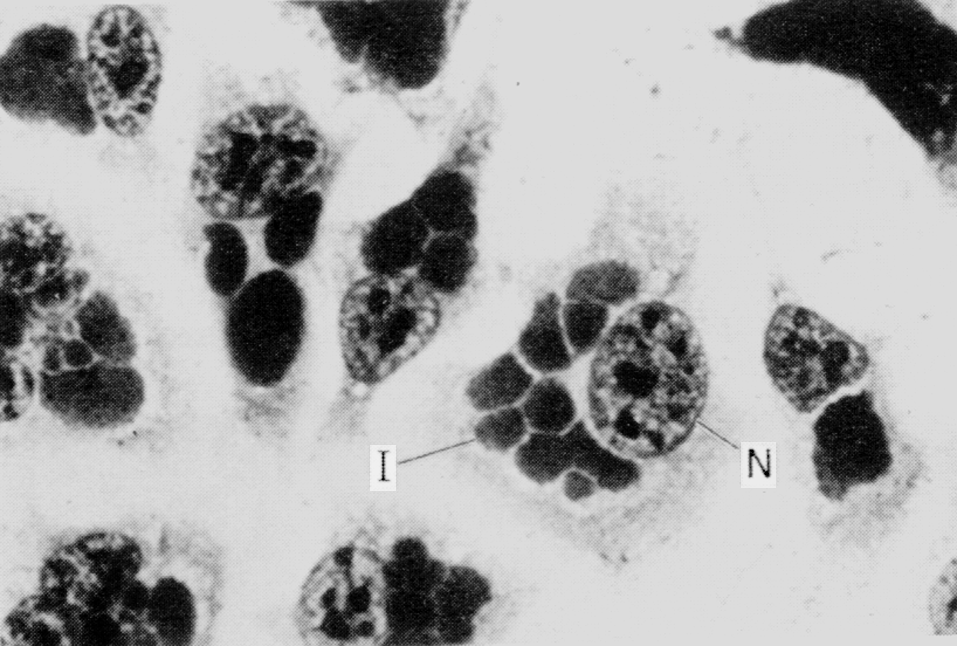

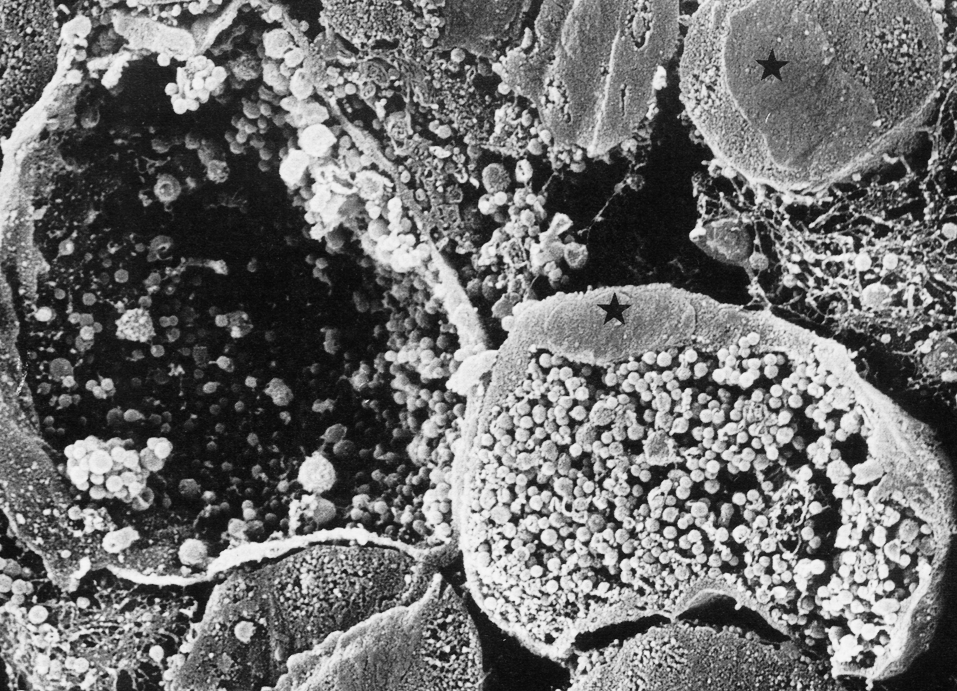
В конце цикла включения (по сути микроколонии хламидий) занимают большую часть клетки и содержат многочисленное потомство ЭТ (рис. 4). Растягивая стенку вакуоли, они разрывают ее, а затем и плазматическую мембрану. Цикл продолжается 40—48 ч, завершаясь гибелью клетки и выходом ЭТ на свободу (рис. 5).

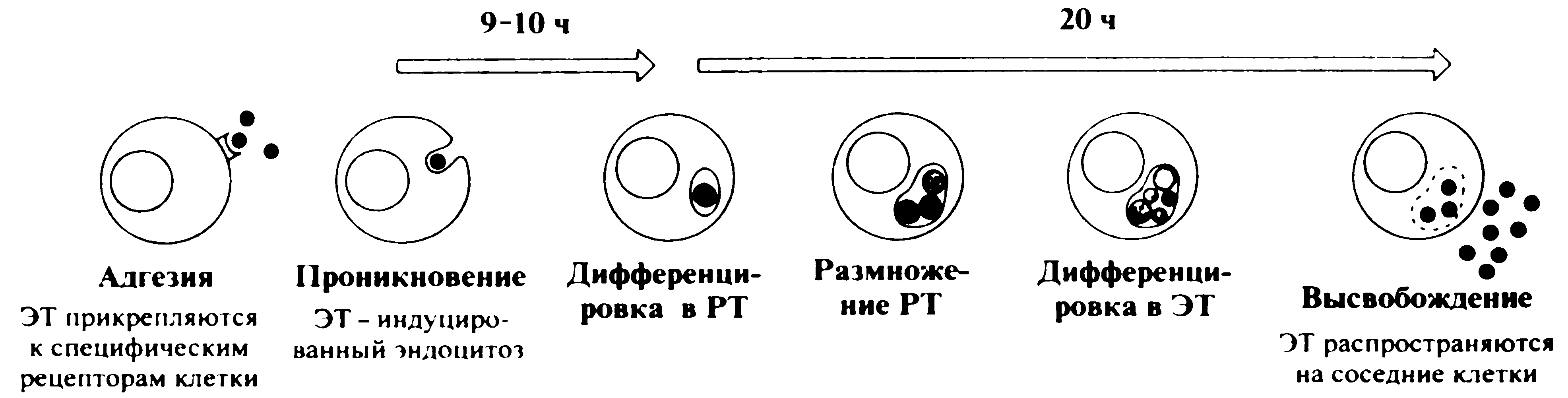
Этому способствует и ослабление конструктивного метаболизма зараженных клеток, которые переключаются на обслуживание бурно реплицирующихся хламидий. В целом это логично объясняет деструктивный эффект хламидий, тем более что никаких специальных цитотоксинов у них не обнаружено. Не следует, впрочем, забывать о том, что при высокой инфицирующей дозе хламидии способны убивать клетки до репликации, т.е. перед тем, как возникает давление внутриклеточных включений, переполненных микробными частицами.
Репликативный цикл может быть искусственно растянут и даже приостановлен, если зараженные клетки поместить в неблагоприятные условия (например, лишить необходимых факторов питания) или добавить ингибиторы размножения хламидий (например, пенициллин). Хламидии не погибают, сохраняя способность к размножению после устранения негативных факторов. В известной мере это моделирует склонность хламидий к латентным (персистентным) инфекциям (см. ниже).
Классификация хламидий формально проста. Порядок Chlamydiales включает одно семейство (Chlamydiaсeae) c единственным родом (Chlamydia). Число видовых таксонов тоже весьма ограничено. До недавнего времени их было всего два — Ch. trachomatis и Ch. psittaci, в 1989 г. введен еще один вид — Ch. pneumoniae. О таксономической самостоятельности каждого из них говорит множество эколого-патогенетических, микробиологических и иммунохимических вариантов. Этому соответствует и слабое генетическое родство видовых таксонов хламидий — всего 5—10% гомологии ДНК.
В 1999 г. предложена новая классификация, привыкание к которой (если оно состоится) потребует времени. Классификация основана на генетических критериях, использующих степень гомологии в нуклеотидной последовательности генов 16S и 23S рибосомальных РНК: >95% для всех представителей одного рода, >90% — семейства и >80% — порядка/класса микроорганизмов. В соответствии с этим семейство Chlamydiaceae разделено на два рода: Chlamydia и Chlamydophila. В состав рода Chlamydia кроме Chlamydia trachomatis вошли Chlamydia muridarum и Chlamydia suis (патогены животных). Род Сhlamydophila составляют Chlamydophila pneumoniae (прежнее название Chlamydia pneumoniae) и варианты прежнего вида Chlamydia psittaci (Chlamydophila psittaci, Chlamydophila abortus, Chlamydophila felis, Chlamydophila pecorum и Chlamydophila caviae). Chlamydophila psittaci включает штаммы 8 сероваров, для которых основными хозяевами являются птицы. Все они могут передаваться человеку, вызывая пситтакоз (см. ниже).
Chlamydia trachomatis
Взаимоотношения с человеком, единственным естественным хозяином Ch. trachomatis (исключение составляют биовары Ch. muridarum и Ch. suis, вызывающие пневмонию у мышей и свиней), определяются способностью данного вида персистировать в цилиндрическом эпителии генитального тракта и конъюнктивы. Часто это протекает бессимптомно, однако в ряде случаев слабое, но рецидивирующее повреждение эпителия возбуждает и поддерживает воспалительную реакцию слизистых оболочек — основу хронической патологии, типичной для Ch. trachomatis.
Принято выделять три разновидности/биогруппы Ch. trachomatis: глазные, генитальные и хламидии венерической лимфогранулемы (ВЛГ). Последние обладают повышенной вирулентностью и способны выходить за пределы слизистых оболочек, поражая регионарные лимфатические узлы. Глазные и генитальные хламидии являются строгими эпителиальными паразитами, лишенными способности инвазировать глубжележащие ткани.
Эколого-патогенетическая неоднородность Ch. trachomatis обрела реальную основу в 1970-х гг., когда при помощи микроиммунофлюоресценции удалось серотипировать Ch. trachomatis. В настоящее время известно 18 cероваров, принадлежность к которым удивительным образом соответствует патогенетическим потенциям хламидий. Серовары А, В, Ва, С включают глазные хламидии, вызывающие трахому; D—K-хламидии служат причиной урогенитального хламидиоза и его экстрагенитальных осложнений; L-серовары (L1, L2, L3) являются возбудителями ВЛГ.
Трахома — это хронический конъюнктивит, в основе которого лежит гранулематозное воспаление конъюнктивы, захватывающее роговицу (кератоконъюнктивит). Начавшись в раннем детстве, заболевание прогрессирует на протяжении многих лет и полностью купируется лишь после зарастания очагов инфекции рубцовой соединительной тканью. Это тот случай, когда микробиологическое выздоровление дается ценой тяжелых структурных изменений, ведущих к потере функции, в данном случае зрения. Эпителий конъюнктивы человека — единственный природный резервуар возбудителя трахомы, его входные ворота и зона патогенетически значимого размножения. Трагический парадокс заключается в том, что сугубо местный процесс, который на первых порах доставляет мало беспокойств и вполне излечим, обретает катастрофическую необратимость. Впрочем, как и при других хламидиозах, процесс нередко ограничивается начальными проявлениями, не достигая тяжких последствий. Более того, возможны бессимптомное инфицирование и даже длительная персистенция без клинических проявлений.
Название болезни пришло из глубокой древности. В переводе с греческого «trachys» означает «шероховатый, неровный». Это связано с тем, что поверхность конъюнктивы выглядит бугристой из-за лимфоидных фолликулов (трахоматозных гранулем), которые формируются в субэпителиальной ткани. В запущенных случаях вся конъюнктива сплошь усеяна зернышками, тесно прилегающими друг к другу, так что получается картина вареного саго. Отсюда об активной фазе болезни говорят как о фолликулярном конъюнктивите, подчеркивая патогенетическую уникальность трахомы в сравнении с другими конъюнктивитами (кстати, одно из старых названий трахомы — зернистое воспаление глаз).
Инфекция не обеспечивает иммунитета. Напротив, иммунный ответ является одним из механизмов болезнетворности хламидий (это относится и к генитальному хламидиозу — см. ниже). На территориях, эндемичных по трахоме, процесс поддерживается вследствие реинфицирования, причем повторное заражение вызывает более сильную реакцию за счет гиперчувствительности, которую оставляют предыдущие контакты с хламидийными антигенами. Выявлено, что к числу таких факторов относится аллергия замедленного типа против белка теплового шока (hsp60). Он активно продуцируется Ch. trachomatis, поддерживая иммунопатогенез хламидийной персистенции.
Большое влияние оказывает вторичная бактериальная инфекция. Она вносит элементы пиогенного воспаления, способствуя деструкции эпителия и развитию реактивного склероза. Среди бактерий доминируют представители нормальной микрофлоры конъюнктивы: гемофилы, моракселлы, пневмококки и др. Они безвредны для неповрежденного эпителия, но ассоциация с хламидиями раскрывает их патогенетический потенциал. Неоднородность микробного пейзажа при трахоме долго (начиная с Р. Коха, заподозрившего в этиологии трахомы обнаруженный им вид гемофилов — Haemophilus aegiptius) вводила в заблуждение, и даже после открытия внутриэпителиальных хламидийных включений скептики продолжали сомневаться в моноэтиологичности трахомы, ссылаясь на недостаточно строгое соблюдение постулатов Коха. Точки над «i» были расставлены лишь в конце 1950-х гг., когда путем заражения хламидиями, изолированными от больных трахомой, удалось воспроизвести болезнь у добровольцев. Это утвердило первичную патогенность Сh. trachomatis для здоровой конъюнктивы. Ее лишены другие бактерии, которые сопутствуют хламидиям в качестве оппортунистических патогенов.
Трахома отличается эндемичностью, даже гиперэндемичностью. Ее родиной являются страны Северной Африки, Ближнего и Среднего Востока, Юго-Восточной Азии и Центральной Америки. Именно в этих регионах сконцентрировано большинство случаев современной трахомы. В Европе болезнь получила распространение после вояжей крестоносцев и особенно в эпоху наполеоновских войн. Ее принесли французские солдаты, заразившиеся во время египетского похода (поэтому трахому называли «египетским воспалением глаз»). Инфекция передается через руки и предметы, загрязненные выделениями из больного глаза (полотенца, носовые платки, посуда для умывания); распространению заразы способствуют мухи.
Из этого следует, что трахома — один из символов низкого уровня санитарной культуры и общественного благополучия. Она всегда доминировала среди беднейших слоев населения, где благоприятными факторами для ее распространения являются скученность и нечистоплотность. В настоящее время в развитых странах трахома встречается в виде спорадических случаев, но в своих «родных» регионах она является большой проблемой, оставаясь самой важной глазной инфекцией. Более 600 млн. людей заражены Ch. trachomatis, из них 10—20 млн. — слепых, многие страдают пороками зрения.
Несмотря на многолетние изыскания, создать вакцину против трахомы (как и для других хламидийных инфекций) не удалось, хотя в культурах in vitro антитела против типоспецифических антигенов нейтрализуют инфекциозность хламидий. Впрочем, все может решить элементарное соблюдение правил гигиены. Например, согласно исследованиям, проведенным в Мексике, ежедневное умывание почти в 5 раз снижает заболеваемость трахомой среди детей из групп риска.
Генитальный хламидиоз. Нынешняя популярность хламидий связана с менее манифестной, но гораздо более распространенной, чем трахома, патологией — поражением урогенитального тракта D—К-сероварами Ch. trachomatis. Значение Ch. trachomatis в этиологии заболеваний, передаваемых половым путем, подчеркивается подразделением негонококковых инфекций (иногда их неудачно называют неспецифическими) на хламидийные и прочие, не ассоциированные с Ch. trachomatis. Первые составляют более половины такого рода заболеваний. Полагают, что ежегодно возникает не менее 50 млн. свежих случаев генитального хламидиоза, а по подсчетам американских авторов, годовые затраты в США, связанные с обслуживанием таких инфекций, превышают 2,4 млрд. долл.
Стабилизация инфекта чаще всего происходит в цилиндрическом эпителии шейки матки, и именно эндоцервикс следует считать основным резервуаром генитальных хламидий, где они длительно и нередко бессимптомно персистируют. Вторым по значению биотопом является уретра (женская и мужская). Изредка инфицируется прямая кишка.
Полагают, что в среднем заражено более 10% женщин, а среди тех, кто обращается за консультацией к венерологам, уровень инфицированности достигает 40%. Клинические проявления женского хламидиоза включают цервицит, уретрит, бартолинит, эндометрит, сальпингит, перигепатит. Но чаще (в 70—80% случаев) инфекция протекает бессимптомно, хотя это не означает полной безвредности для носителя, не говоря об эпидемиологической опасности, т.е. передачи инфекции окружающим. Главные проблемы связаны с восходящей инфекцией, так называемыми воспалительными заболеваниями малого таза. Более половины случаев хронического (часто молчащего) сальпингита ассоциировано с хламидиями. Финалом является трубная непроходимость со всеми вытекающими отсюда последствиями — от эктопической (внематочной) беременности до бесплодия. Менее очевидным выглядит влияние Ch. trachomatis на течение беременности. Сообщения о более высоком проценте мертворождения и преждевременных родов среди инфицированных хламидиями женщин не нашли подтверждения.
У мужчин Ch. trachomatis первично инфицирует эпителий уретры. Примерно в половине случаев это проходит бессимптомно (хламидии обнаруживаются у 4—10% здоровых мужчин), в остальных случаях через 1—3 нед после полового контакта развивается уретрит, протекающий менее остро, чем гонорея. Хламидийные уретриты составляют 30—60% случаев негонококкового уретрита — самого распространенного заболевания мужчин, передаваемого половым путем. Нередко именно мужские уретриты манифестируют латентную хламидийную инфекцию женщин. Даже тогда, когда хламидийный уретрит протекает субклинически, он может осложниться синдромом Рейтера (см. ниже) или эпидидимитом, т.е. воспалением семенных протоков. Последнее чревато потерей способности к оплодотворению и наряду с сальпингитом служит одной из причин бесплодных браков. Хламидии не склонны длительно персистировать в уретре, но благодаря половым контактам получают отличную возможность для распространения.
Сегодня мало кто сомневается в том, что Ch. trachomatis может поражать здоровый эпителий слизистой оболочки урогенитального тракта, т.е. выступать в роли первичного патогена. Но ясно и то, что агрессивность хламидий возрастает при смешанной инфекции, прежде всего в связке с гонореей. Они нередко передаются вместе с гонококками и, выживая в условиях антимикробной терапии, обеспечивают хронизацию процесса. Благодаря современным антибиотикам это наблюдается реже, но в эпоху пенициллинотерапии было обычным явлением. Например, постгонококковые уретриты наблюдались у 1/4—1/3 больных и в 40—90% cлучаев были ассоциированы с Ch. trachomatis. Не исключено, что гонококки активируют законсервированные в эпителиоцитах хламидии, но проще допустить, что гонорейный процесс и, возможно, другие инфекции повышают вероятность колонизации эпителиоцитов экзогенными хламидиями.
Клинически урогенитальные хламидийные инфекции не отличимы от таких же заболеваний иной этиологии. Характерным для патогистологической картины являются лимфоидные микрофолликулы в пораженных участках слизистой оболочки. Они напоминают трахоматозные зерна при хламидийных конъюнктивитах и имеют, по-видимому, отношение к патогенезу (иммунопатогенезу?!) всех хламидиозов (см. выше).
Подобно трахоме, генитальный хламидиоз оставляет антитела, но не оставляет иммунитета, или он слишком специфичен, чтобы защитить от гетерологичных штаммов Ch. trachomatis. Поэтому многие больные в короткие сроки инфицируются повторно, причем у женщин вероятность осложнений (таких, как внематочная беременность, бесплодие) повышается с каждым эпизодом реинфицирования.
Конъюнктивит. Родство генитальных и глазных хламидий проявляется в том, что D—К-разновидности Ch. trachomatis cпособны вызывать у человека острый конъюнктивит, который персистирует в течение месяцев и патоморфологически (фолликулярный конъюнктивит с внутриэпителиальными включениями) похож на трахому. Инфицирование глаз происходит в результате прямого контакта с материалом из половых органов. Не случайно урогенитальный тракт (шейка матки или уретра) у 90% женщин и мужчин, страдающих конъюнктивитом с включениями (официальное название хламидийной инфекции глаз), заражен Ch. trachomatis. Так же, как генитальный хламидиоз, это часто является следствием беспорядочных половых связей. Описаны и случайные заражения при попадании в глаза инфекционного материала, например у гинекологов. Замечено, что конъюнктивиты с включениями редко бывают у гомосексуалистов, чему соответствует и низкий уровень хламидийных поражений уретры в этой группе лиц. Неясным остается отношение хламидий к конъюнктивитам плавательных бассейнов, или бассейновым конъюнктивитам. В ряде случаев их вызывают аденовирусы, заражение которыми может также происходить в плавательных бассейнах.
В отличие от трахомы, конъюнктивит с включениями встречается повсеместно, протекает менее упорно и не столь тяжело. Без лечения обычно через несколько месяцев наступает самоизлечение. Впрочем, поручиться за это нельзя, так как, подобно трахоме, процесс изредка ведет к необратимым (рубцовым) изменениям конъюнктивы и роговицы. Отсюда и пугающее название — паратрахома, которое с современных позиций следует воспринимать как не более чем формальное сходство с классической трахомой и ее возбудителем.
Синдром Рейтера. Сh. trachomatis причастны еще к одной, очень коварной патологии — синдрому Рейтера. В классическом варианте он сочетает в себе триаду признаков: уретрит — конъюнктивит — артрит (уретроокулосиновиальный синдром), к которым часто добавляются полиморфные поражения кожи и слизистых оболочек. Впервые описанный как хроническое потенциально инвалидизирующее осложнение кишечных инфекций (шигеллы, сальмонеллы, йерсинии, кампилобактер), синдром Рейтера был распространен и на заболевания, передаваемые половым путем, прежде всего генитальный хламидиоз. Независимо от этиологии (кишечная или генитальная формы) синдром развивается как реактивная (т.е. напрямую не связанная с действием живого возбудителя) патология через несколько недель после острой инфекции, которая может остаться незамеченной или не привлечь к себе серьезного внимания (больной не обращается к врачу). Классическая картина наблюдается не всегда: возможны разные сочетания симптомов. Ведущим является поражение суставов (как правило, синовиты нижних конечностей), и понятие «реактивный артрит» иногда вычленяется в самостоятельную нозологическую категорию, хотя с этиопатогенетической точки зрения это не более, чем вариант синдрома Рейтера, как и другие постинфекционные осложнения реактивной природы.
О причинах развития синдрома почти ничего не известно. Рискуя повториться (см. «Йерсинии»), заметим, что он в 10 раз чаще наблюдается у мужчин (особенно молодого возраста, 20—30 лет) и зависит от генетического фона: 65—75% заболевших имеют антиген гистосовместимости HLA-B27. Артриты возникают в течение месяца примерно у 1% мужчин с негонококковым уретритом; для носителей HLB-B27 вероятность достигает 20%. Полная триада Рейтера наблюдается в 30—50% случаев. Присутствие живых хламидий в синовиальных оболочках (как и в других очагах поражения) необязательно, хотя такие находки и удавались. С большим постоянством в них обнаруживаются хламидийные антигены, которые, возможно, запускают и поддерживают иммунологические механизмы повреждения у генетически предрасположенных лиц. На основании этого для лечения синдрома Рейтера предлагается длительное (в течение двух и более месяцев) лечение антибиотиками, чтобы искоренить генитальные хламидии. Но скорее всего это формальная, а потому вредная логика. Не исключено, что микробные антигены выполняют лишь триггерную функцию и, возбудив реакцию, «покидают сцену». По крайней мере ни артриты, ни кожные и глазные поражения при этом синдроме не поддаются действию антибиотиков, и многие сомневаются в их эффективности. В связи с этим главное (но, увы, не лучшее) лечение сводится к симптоматической борьбе с асептическим воспалением (нестероидные противовоспалительные средства, реже кортикостероиды). По сути полагаются на жребий, ибо при статистически высокой вероятности выздоровления судьба каждого больного непредсказуема. По приблизительным оценкам около половины полностью выздоравливают после первой и единственной атаки. Из второй половины часть страдает редкими рецидивами, у других развивается хронический артрит и прогрессирующий спондилит. Если не выдавать желаемое за действительное, приходится констатировать, что лекарства против синдрома Рейтера не существует.
Своеобразие синдрома Рейтера, неразгаданность его патогенеза (а возможно, и истинных этиологических факторов) рождает немало спекуляций, в основном иммунопатологического толка. Например, у хламидий обнаружена способность к поликлональной стимуляции В-лимфоцитов, которая ведет к образованию широкого спектра случайных антител, в том числе аутоантител к различным тканям человека. Однако такого рода В-суперантигенность наблюдается и у других бактерий, не причастных к развитию синдрома Рейтера. Более того, при хламидиозах гипергаммаглобулинемия (результат поликлональной стимуляции В-лимфоцитов) далеко не всегда осложняется реактивной патологией.
Инфекции новорожденных. Присутствие хламидий в цервикальном канале беременных женщин (его вероятность в среднем оценивается как 12%) представляет опасность для новорожденных. Заражение происходит в родовом канале, и уже в первые дни жизни примерно 65% детей, родившихся от инфицированных матерей, колонизированы хламидиями. По данным зарубежных коллег, 28 из 1000 новорожденных контаминированы хламидиями. О том, что это не индифферентная встреча, говорит сероконверсия (т.е. образование антител против хламидий), которая наблюдается в 70% случаев. Хламидии удается выделить из конъюнктивы, носоглотки, среднего уха, трахеи, легких, прямой кишки, влагалища. Это означает, что число неонатальных синдромов, предположительно связанных с хламидиями, может быть достаточно велико, хотя реальное значение имеют лишь инфекции глаз и легких. В первом случае возникает конъюнктивит (бленорея новорожденных), во втором — атипичная пневмония. Риск развития конъюнктивита у детей, родившихся от матерей с хламидийной инфекцией шейки матки, составляет 25%, воспаления легких — 10%. Четвертая часть детей до 6 мес, госпитализируемых по поводу патологии нижних отделов респираторного тракта, и три четверти детей с афебрильной пневмонией заражены хламидиями. Принято считать, что хламидии первично инфицируют конъюнктиву и уже затем через слезный канал спускаются в респираторный тракт. Но это необязательно, и примерно у половины детей пневмония возникает независимо от бленореи.
Степень клинических проявлений колеблется в широких пределах — от стертых до угрожающе манифестных форм. Процесс продолжается несколько недель и даже месяцев и, как правило, заканчивается выздоровлением. Но возможны осложнения, например рубцевание конъюнктивы и поверхностная васкуляризация (паннус) роговицы, типичные для классической трахомы. Пневмония иногда надолго делает легкие функционально неполноценными.
В конъюнктивальных мазках обнаруживаются внутриэпителиальные включения, типичные для Ch. trachomatis (тельца Гальбштедтера—Провацека; см. рис. 1). Это один из признаков, по которым хламидийная офтальмия (конъюнктивит с включениями) отличается от гонококковой бленореи. Закапывание в конъюнктивальный мешок 1% раствора нитрата серебра, практикуемое с конца прошлого века для профилактики гонококковой инфекции у новорожденных, не предупреждает развития хламидийного конъюнктивита, распространенность которого заметно выше, чем гонококковой бленореи. Это побуждает к использованию иных профилактических средств, в частности местной аппликации антибиотиков.
При высоком уровне хламидиоза среди новорожденных целесообразно обследовать беременных женщин с целью выявления и элиминации хламидий. Особое внимание следует обращать на активные формы генитального хламидиоза, которые сопровождаются воспалительной реакцией. Именно в таких случаях концентрация хламидий в зоне инфекции особенно высока, а потому эпидемически опасна. В целом по распространенности врожденный хламидиоз можно сравнить с верхушкой айсберга, основой которого служат урогенитальные инфекции взрослых.
Венерическая лимфогранулема. Это заболевание характерно для стран тропического и субтропического поясов, хотя единичные случаи встречаются повсеместно. ВЛГ-хламидии (L-cеровары) отличаются повышенной инвазивностью и, первично размножаясь в эпителии слизистой оболочки полового тракта и/или прямой кишки, внедряются и реплицируются в регионарных (обычно паховых) лимфатических узлах. Важнейшими признаками ВЛГ являются местные лимфадениты, гнойные фистулы, проктиты, абсцессы и рубцовые поражения прямой кишки. Мужчины болеют чаще и страдают сильнее, чем женщины (вероятно, из-за особенностей лимфатической системы половых органов).
Памятуя об истории открытия хламидий, можно, казалось бы, не опасаться за диагностику хламидийных инфекций: очевидным представляется выявление клеток с характерными включениями. Но дело обстоит сложнее. Прямой цитологический метод пригоден лишь для констатации конъюнктивита новорожденных (чувствительность более 90%) и малоинформативен при хламидийных (особенно, генитальных) инфекциях у взрослых.
В 1994 г. Рокфеллеровский фонд (США) предложил 1 млн. долларов за быстрый и надежный тест для диагностики генитального хламидиоза и гонореи, применимый в экономически отсталых странах. Эта мечта до сих пор не реализована. Единственным надежным способом выявления C. trachomatis до недавнего времени оставался культуральный метод — выделение хламидий из исследуемого материала в культуре клеток. Трудоемкость и высокая стоимость процедуры сдерживали широкомасштабные исследования. Ситуация изменилась в 1980-х гг., когда были разработаны высокочувствительные некультуральные методы обнаружения хламидий. Они основаны на выявлении антигенов (иммунофлюоресцентная микроскопия, иммуноферментный анализ) и нуклеиновых кислот (гибридизация ДНК, полимеразная цепная реакция и др.) и по чувствительности приближаются к «золотому стандарту» (выделению чистой культуры) или превосходят его. Однако трактовка результатов (прежде всего позитивных) требует деликатности. Их следует оценивать с учетом анамнеза, активности процесса и по возможности подтверждать другими методами.
Серологические тесты (определение антихламидийных антител в крови и секрете слизистых оболочек) не гарантируют диагноза свежей инфекции. Это связано со слабой антигенной нагрузкой при местном вялотекущем процессе и широкими контактами с хламидиями. В любом случае ни обнаружение антител (у многих они имеются до очередного заражения), ни сероконверсия (для достоверного повышения титра антител может потребоваться месяц и более), ни попытки выявления IgM-антител (IgM-фаза отсутствует вследствие повторных контактов с возбудителем) не решают проблемы. Отметим, впрочем, два исключения: диагностическая значимость IgM-теста при хламидийной пневмонии новорожденных и сероконверсия у больных ВЛГ. В каждом из этих случаев речь идет о системном процессе, а потому о более интенсивном и быстром иммунном ответе, чем при локальной инфекции генитального тракта. Кроме того, новорожденные, которые впервые взаимодействуют с C. trachomatis, отвечают опережающим образованием IgM-антител.
При лечении генитальных хламидиозов следует учитывать характер заболевания: острые или хронические инфекции, связанные с малосимптомной латенцией возбудителя. Если в первом случае задача решается успешно (например, купирование острого уретрита), то воздействие на персистирующие инфекции сопряжено с серьезными трудностями. Это объясняется, как минимум, двумя причинами — низкой репликативной активностью хламидий и высокой выживаемостью элементарных тел. Для усиления эффективности предложена цикловая терапия, которая сочетает в себе несколько последовательных курсов лечения антибиотиками с короткими перерывами. Расчет строится на уничтожении вегетативных форм (ретикулярных тел), очередном прорастании элементарных тел и новых атаках на возникшие из них вегетативные клетки. Словом, все, как при стерилизации (тиндализации) пищевых продуктов для избавления от спорообразующих бактерий.
Неоднозначен и вопрос о микробиологических критериях выздоровления. Тесты на хламидийные антигены и ДНК нередко остаются позитивными после лечения, не говоря о таком консервативном индикаторе, как антитела. Есть мнение, что опорой должно быть отсутствие живых хламидий, т.е. негативный результат бактериологического (культурального) анализа. Однако сделать это для хламидий непросто, а в рутинном варианте невозможно. Приходится ориентироваться на отсроченные (спустя 1—1,5 мес после лечения) поиски маркеров, чтобы дождаться уничтожения пассивных следов инфекции. Но за этот срок может произойти реинфицирование, и все надо начинать сначала.
Сhlamydia pneumoniae
История данного вида хламидий началась на Тайване в 1965 г., когда от ребенка с трахомой был изолирован атипичный хламидийный штамм. В 1983 г. такой же штамм выделили в США от больного фарингитом. Все последующие изоляты были получены от больных с заболеваниями респираторного тракта (пневмония, бронхит, синусит, фарингит), среди которых было выявлено немало лиц с антителами против вновь обнаруженных хламидий. Последние получили рабочий акроним TWAR от начальных букв первых изолятов — Taiwan (TW) и acute respiratory (AR) infection. Это обозначение в ходу до сих пор даже после того, как в 1989 г. Grayston предложил современное название — Chlamydia pneumoniae, подчеркивающее важность данной разновидности хламидий в патологии легких человека.
Ch. pneumoniae имеет глобальное распространение. Удивительным оказалось то, что, в отличие от внутривидовой гетерогенности Ch. trachomatis и Сh. psittaci, различные изоляты Ch. pneumoniae (в том числе выделенные в разных регионах Земного шара) не отличимы друг от друга. Они антигенно идентичны, не отличаются по рестриктазным фрагментам ДНК, структуре главного белка наружной мембраны и обладают 94—100% ДНК-гомологией. Словом, есть веские основания для необычного вывода: все известные на сегодняшний день изоляты Ch. pneumoniae являются близнецами штамма TWAR, открывшего новую страницу хламидологии.
Таксономическую самостоятельность Ch. pneumoniae подчеркивает наличие видоспецифического антигена и слабое генетическое родство (5—10% гомологии ДНК) с другими хламидиями. Кроме того, элементарные тела Сh. pneumoniae имеют не шаровидную, а копьевидную форму, что может быть важным для развития инфекции, так как именно заостренным концом эти тела прикрепляются к эпителиальным клеткам.
Единственный резервуар инфекции — человек. Согласно данным серологического анализа, контакт с Ch. pneumoniae наблюдается чаще, чем с другими хламидиями: 60—80% взрослого населения имеет специфические антитела. Они редко обнаруживаются у детей дошкольного возраста, быстро нарастают к 20 годам и, появившись, сохраняются на всю жизнь, по-видимому, благодаря периодическому реинфицированию, которое внешне никак не проявляется. В целом иммунологически манифестные (об этом судят по нарастанию титра антител), но клинически не выраженные или малосимптомные формы достаточно типичны для заражения Ch. pneumoniae.
Бронхит и пневмония наблюдаются в небольшом числе случаев, чаще после 20 лет. Однако, если учитывать широкое распространение Ch. pneumoniae, патология глубоких отделов респираторного тракта — вовсе не редкость. Ch. pneumoniae считается причиной около 10% случаев пневмонии и по частоте занимает 3—4-е место среди ее других возбудителей. Заболевания наблюдаются в виде отдельных случаев и вспышек, которые наслаиваются на спорадическую заболеваемость. Возбудитель передается скорее всего воздушно-капельным путем, но распространение инфекции идет медленно: манифестные формы появляются с интервалом в среднем 30 дней. Это можно объяснить тем, что многие больные выделяют слишком мало хламидий, чтобы заразить окружающих. Не исключено, что эпидемически значимым источником служат промежуточные носители, которые бессимптомно «подкапливают» Ch. pneumoniae в верхних отделах респираторного тракта.
Патология легких, связанная с Ch. pneumoniae, принадлежит к атипичным пневмониям и клинически не отличима от других инфекций подобного типа (Mycoplasma pneumoniae, Legionella pneumophila, респираторные вирусы). Начало — вялое, лейкоцитоза, как правило, не бывает, температура повышается умеренно и к моменту обращения к врачу может отсутствовать. Но даже при внешне бедной симптоматике хрипы в легких прослушиваются регулярно. Нередко одной из первых жалоб или отсроченных осложнений является синусит.
В большинстве случаев заболевание протекает сравнительно легко. Так же, как микоплазменную пневмонию, многие больные переносят его на ногах и не требуют госпитализации. Однако выздоровление нередко идет медленно (кашель и слабость иногда сохраняются несколько недель и даже месяцев), и даже несмотря на прицельную антибиотикотерапию (тетрациклины, макролиды), возможно длительное бактерионосительство. Более того, изредка наблюдаются тяжелые осложнения вплоть до фатального исхода. Обычно это происходит на фоне другой патологии или связано с вторичными осложнениями, например пневмококковой бактериемией.
Неожиданная интрига возникла после сообщений о том, что антитела к TWAR/Ch. pneumoniae ассоциированы с хронической коронарной болезнью и острым инфарктом миокарда. Сероэпидемиологические исследования подтвердили корреляцию между содержанием антихламидийных антител и атеросклеротическим поражением коронарных сосудов. Прицельные поиски позволили обнаружить Ch. pneumoniae в атероматозных бляшках коронарных артерий и аорты, но не в нормальных сосудах. Это вызывает большой интерес, но вместе с тем требует сдержанной интерпретации. Гипотеза об инфекционной природе атеросклероза, который сегодня рассматривается как воспалительная болезнь, не нова и, безусловно, выходит за рамки хламидиозов. Словом, до профилактики инфаркта миокарда антибиотиками еще далеко, и хочется верить, что увлечения не перейдут разумных границ. Феномен присутствия хламидий в зоне атеросклеротических изменений может быть вторичным, хотя, не исключено, что инфекция поддерживает процесс. В любом случае проникновение в стенку сосудов говорит о повышенной инвазивности Ch. pneumoniae и возможности дистантных осложнений при поражениях респираторного тракта.
Перенесенное заболевание не оставляет прочного иммунитета. По крайней мере, устойчивость к реинфицированию не зависит от сывороточных антител. Известны случаи повторной пневмонии и более легких проявлений инфекции. В этом отношении Ch. pneumoniae повторяет другие хламидии, хотя, если принять во внимание ее антигенную монотонность, здесь это вызывает больше вопросов.
Для лабораторного подтверждения диагноза можно воспользоваться определением специфических антител, т.е. иммуносеродиагностикой. С этой целью разработаны иммуноферментные тест-системы для опредления видоспецифических антител классов G, A и M. Наличие IgM-антител подтверждает острую или недавнюю инфекцию, продолжительное присутствие короткоживущих IgA-антител считается индикатором хронической инфекции. Что касается IgG-антител, то только их значительное нарастание в парных сыворотках дает возможность предполагать наличие Ch. pneumoniae. Надежность выводов уменьшается при повторных инфекциях (обычно у взрослых), так как в таких случаях отсутствуют IgM-антитела. Этим можно объяснить и низкую чувствительность реакции связывания комплемента, которая во многом зависит от IgM-антител: лишь 10—25% больных имеют диагностически значимые титры комплементсвязывающих антител.
Выделение Ch. pneumoniae сопряжено с еще бульшими трудностями, чем других хламидий, так как мокрота, из которой пытаются выделить возбудителя, нередко вызывает гибель клеток. Кстати, клеточные культуры различаются по чувствительности к хламидиям: Ch. pneumoniae размножается в клетках линий HL и Нер-2, Ch. trachomatis — HeLa и МсСoy, а Сh. psittaci непривередливы в этом отношении и реплицируются в большинстве клеточных культур.
Ценным диагностическим критерием является ПЦР-анализ. Есть данные, что чувствительность ПЦР при выявлении Сh. pneumoniae из образцов назофарингеальных смывов на 25% превосходит культуральный метод. Это означает, что ПЦР может стать новым «золотым стандартом» в диагностике этой инфекции.
Выявление микробных антигенов не нашло широкого применения из-за низкой чувствительности и специфичности.
CHlamydia psittaci
В отличие от Ch. trachomatis и Ch. pneumoniae эти хламидии занимают скромное место в патологии человека. Причина в том, что они паразитируют на животных (птицах и млекопитающих), а человек выступает в роли случайного хозяина, изредка заражаясь от птиц. Животные обычно мирно уживаются с Ch. psittaci, но иногда хламидии активируются и вызывают тяжелые заболевания: поражения респираторного тракта, выкидыши, полиартриты, энтероколиты, конъюнктивиты, энцефаломиелиты и пр. Болезни домашних животных (птица, крупный и мелкий рогатый скот) приносят ощутимый экономический ущерб, в связи с чем хламидиозы служат предметом серьезного внимания ветеринарных микробиологов.
Широкое распространение в природе (инфицированы едва ли не все виды птиц и млекопитающих, за исключением человека и приматов) заставляет размышлять о причинах столь высокого экологического универсализма Ch. psittaci. Одним из объяснений может быть внутривидовая неоднородность, которая значительно выше, чем у Ch. trachomatis. Ch. psittaci располагает множеством сероваров с различной вирулентностью и широким кругом хозяев. Предложено дифференцировать около десятка биогрупп Ch. psittaci, таких как возбудители выкидыша овец и коров; полиартрита, конъюнктивита и энцефаломиелита овец и коров; перикардита, полиартрита и аборта свиней; пневмонии кошек; конъюнктивита с включениями морских свинок и др. Специальная биогруппа представлена штаммами птиц. Словом, с эколого-патогенетических позиций современный вид Ch. psittaci — достаточно сложное понятие, и в будущем он скорее всего будет разделен на несколько видовых таксонов. Это уже нашло отражение в новой классификации хладимий, где Сhlamydia psittaci представлены пятью видами в новом родовом таксоне Chlamydophila (см. выше).
Для человека штаммы млекопитающих практически не опасны, но он высокочувствителен к птичьим разновидностям. Для птиц (как и для других животных) типична латентная инфекция желудочно-кишечного и реже респираторного трактов. В стрессовых ситуациях (плохие условия содержания, недостаток корма и пр.) хламидии активируются, вызывая поражения органов пищеварения и дыхания, конъюнктивы, ретикулоэндотелиальной системы и ЦНС. Около 30% заболевших птиц погибает в течение 8—9 дней. В активном периоде множество хламидий выделяется с испражнениями и респираторно-конъюнктивальной слизью. После высыхания (это не убивает элементарные тела) они энтеральным и/или респираторным путем заражают других птиц, поддерживая бессимптомную циркуляцию в природе, а иногда и эпизоотический процесс. Для некоторых видов птиц известна вертикальная (трансовариальная) передача инфекции.
Из сказанного следуют два важных вывода. Во-первых, благодаря способности реплицироваться в разных типах клеток Ch. psittaci потенциально опасны для различных систем организма, впрочем, достаточно и того, что они размножаются в макрофагах, которые представлены во всех тканях, создавая универсальный плацдарм для паразитирующих в них микробов. Второй вывод касается высокой инвазивности Ch. psittaci. У заболевших животных они проникают в кровь, а потому ассоциированные с ними процессы обретают генерализованный характер, хотя и зависят от входных ворот инфекции.
Заболевание, которое Ch. psittaci вызывает у человека, имеет два равноценных названия: пситтакоз и орнитоз. Пситтакоз означает по-гречески «попугай» и отражает давно подмеченную связь между пневмонией и контактом с попугаями. Но лишь после вспышки в 1929—1930 гг. (в Латинской Америке и Европе заболело около 600 человек) пситтакоз получил статус самостоятельной нозологической единицы, а его возбудитель признан одним из видов хламидий. Вскоре оказалось, что заразиться можно не только от попугаев, но и от других птиц. Появилась новая болезнь — орнитоз (от лат. ornis — птица), хотя после того, как была доказана общность птичьих штаммов хламидий, понятия «пситтакоз» и «орнитоз» стали синонимами.
Итак, природным резервуаром Ch. psittaci, опасным для человека, являются птицы. Непосредственным источником заражения служат домашние (особенно утки и индюшки), комнатные, дикие водоплавающие птицы, голуби. Возбудитель передается воздушно-пылевым путем при вдыхании пыли, загрязненной выделениями больных птиц. Заболевание обычно имеет профессиональный акцент: болеют работники птицеферм, владельцы декоративных птиц, охотники на промысловую дичь. Случаи передачи инфекции от человека человеку казуистичны и связаны, по-видимому, с особо вирулентными штаммами. В большинстве руководств об этом даже не вспоминается: больной считается незаразным.
Клинические симптомы возникают не у всех инфицированных. Об этом говорит значительная прослойка серопозитивных здоровых лиц среди групп риска по контакту с Ch. psittaci (например, работники птицеферм, продавцы зоомагазинов). Если заболевание все же наступает, оно проявляется в двух формах. Чаще всего возникает атипичная гриппоподобная пневмония с высокой температурой, головной болью, сухим кашлем, мышечными и суставными болями, отсутствием изменений в лейкоформуле или лейкопенией. Другим вариантом является тифоподобная (токсическая) форма: в этом случае признаки тяжелой интоксикации развиваются без респираторного синдрома. Бактериемия в первые дни заболевания определяет возможность диссеминации возбудителя по организму. Вкупе со способностью к репликации в различных типах клеток, включая макрофаги, это создает вероятность осложнений со стороны сердечно-сосудистой системы (миокардит, эндокардит), ЦНС (менингоэнцефалит), печени (гепатит) и др. Тяжелые случаи заканчиваются летальным исходом на фоне кардиоваскулярной и респираторной недостаточности.
Выздоровление затягивается на несколько недель, иногда болезнь принимает хроническое течение с ранними (через 2—4 нед) и поздними (через 3—6 мес) рецидивами. Этому соответствует медленное освобождение от возбудителя, который может длительно и активно персистировать в респираторном тракте после клинического выздоровления. Агрессивность Ch. psittaci вынуждает организм интенсивно реагировать на инвазию. Во всяком случае, иммунологический ответ при пситтакозе выражен сильнее, чем при патологии, связанной с Ch. trachomatis. Впрочем, это тоже не спасает от рецидивов и повторных заболеваний, хотя и помогает в диагностике. Например, здесь легче проследить за нарастанием титров антител: в течение 10—20 дней после начала заболевания это может быть сделано при помощи такого сравнительно малочувствительного метода, как реакция связывания комплемента. При однократном анализе ориентируются на высокие титры (1:64 и более) и наличие IgM-антител.
Окончательно рассеять сомнения можно, выделив возбудителя из крови (первые дни заболевания) или из мокроты (на протяжении всего активного периода). Однако это не только малодоступно, но и опасно. Внутрикожная проба с так называемым орнитозным аллергеном или орнитином, о которой по инерции вспоминают в некоторых руководствах, малоспецифична, а потому ненадежна. Выявление хламидийных антигенов (в том числе в составе внутриклеточных включений) не нашло практического применения. Поэтому, как правило, ограничиваются определением антител. Хорошим подспорьем служит эпидемиологический анамнез (контакт с птицами).
Подобно всем хламидиям, Ch. psittaci наиболее чувствительны к антибиотикам тетрациклинового и макролидного ряда, которые являются препаратами выбора в лечении пситтакоза. Полезно помнить, что сульфаниламиды, действуя на Ch. trachomatis, не влияют на размножение Ch. psittaci.